Trudy M. Wassenaar et al., |
概要 |
| 健康な微生物叢の重要性は強調しすぎることはありません。その組成の乱れは、他の臓器にまで及ぶ可能性のあるさまざまな症状を引き起こす可能性があります。同様に、他の臓器の急性または慢性の状態は、腸内細菌叢の組成と生理機能に影響を与える可能性があります。ここでは、腸と肺の軸に沿った臓器間コミュニケーション、および肺と冠状動脈性心臓病の間、および心血管疾患と腸内細菌叢の間の相互作用について説明します。 COVID-19感染の臨床転帰にも影響を与えるこの器官の三角形は、免疫細胞や短鎖脂肪酸(SCFA)やトリメチルアミン-N-オキシド(TMAO)などの免疫調節因子を含む多数の受容体とエフェクターによって接続されています。腸内細菌叢はこれらのそれぞれにおいて重要な役割を果たし、したがって肺と心臓の健康に影響を及ぼし、この相互作用は両方向で起こります。腸内細菌叢は、プロバイオティクスの経口摂取によって影響を受ける可能性があります。臓器間コミュニケーションの原因となるメカニズムの理解が深まったことで、「理想的な」プロバイオティクスが持つべき要件と、この三角形におけるその役割を定義し始めることができます。 |
1.はじめに |
腸内細菌叢は、個人の全体的な健康にとって重要な要素ですが、さまざまな医学的および心身医学的状態との関係は複雑です。健康な個人間の微生物叢成分のばらつきは広範囲です[1]。この個人間の変動は、食事、遺伝学、微生物群集および感染症への曝露、および投薬を含む多くの要因の結果です[2,3]。微生物叢の構成にはばらつきがありますが、一般的なパターンを認識することができます。これは、個人の健康状態やさまざまな病状と大まかに相関しています。腸内細菌叢は、多くの状態とその治療によって歪む可能性がありますが、微生物叢の歪みもそのような状態の発症または重症度に寄与する可能性があるため、原因と結果を解明することが難しい場合があります。腸内細菌叢は、免疫系および神経内分泌系と常に直接通信しており、栄養素やその他の摂取した化合物を血流に供給することに加えて、迷走神経を介して間接的に脳と情報を交換します。このように、それは一般的な生理学と健康の監視と緩和の両方に関与する主要なプレーヤーです。一時的な歪みを除いて、個々の腸内細菌叢は3歳前後から比較的安定しています[4]。したがって、食事と健康状態が変わらない限り、生涯を通じて、個人の腸内微生物叢の代謝能力はかなり一定のままです。ただし、その組成は、病原体(細菌、真菌、ウイルス、および寄生虫)、意図的または偶発的な抗菌活性を持つ薬物、または非病原性またはプロバイオティクス生物の摂取によって影響を受ける可能性があります。 |
| 腸内細菌叢が心臓血管の健康に大きな影響を与えることがますます明らかになっています。 冠状動脈アテローム性動脈硬化症および関連する心筋障害は、西欧諸国における罹患率および死亡率の主な原因です。 心血管疾患の認識されている危険因子には、高血圧、糖尿病、喫煙、および不利なライフスタイルが含まれます。 ここでは、心血管疾患(CVD)という用語を使用して、アテローム性動脈硬化症とその心不全や血栓症などの結果をまとめて説明します。 不利なライフスタイルには、治療と予防の潜在的なターゲットを提供する食事パターンが含まれます。 腸内細菌叢は、心血管疾患患者と、リスクのある心血管疾患集団では、対照と比較して異なることが示されていますが、原因と結果を常に分離できるとは限りません。 ほとんどの場合、腸内細菌叢と宿主の健康状態は、複数の複雑なインタラクティブな方法で相互に影響を及ぼします。 |
| 腸内細菌叢と肺の健康との相互作用は以前から知られていましたが、COVID-19パンデミックの間に強く再浮上しました[5,6]。 SARS-CoV-2ウイルスは主に上気道に感染しますが、肺に強い合併症があります。軽度から重度のCOVID-19患者の多くでは、腸内細菌叢がひどく乱されており、この乱れは数か月にわたって持続する可能性があります。他の呼吸器ウイルス感染症と同様に、COVID-19は胃腸症状を伴う可能性があり、ウイルスRNAは、感染者の便の症状が終了した後、長期間検出される可能性があります[7]。気道感染の結果としての腸内細菌叢の乱れは、腸内細菌に反応するだけでなくその組成を調節する免疫系および神経系を介して腸内細菌の影響経路が存在するため、必ずしも腸内で活発に複製するウイルスに依存するわけではありません。免疫調節は両方向に働き、肺と腸の微生物叢の両方に影響を及ぼします。健康な腸内微生物叢の組成は、一般的な風邪の感染症で実証されているように、気道感染症とその症状に対するより高い耐性に寄与する可能性があります。 |
| 血液凝固と心臓組織の感染の両方がCOVID-19の一般的な合併症であり、両方ともその死亡率に大きく寄与するため、心血管疾患とCOVID-19(および他のウイルス性肺感染症)との関連も明らかです。 SARS-CoV-2感染は、毛細血管の内皮細胞損傷を間接的に引き起こし、重度のCOVID-19の特徴である多臓器損傷を引き起こす微小血栓症の一因となります[8]。 肥満や糖尿病などの心血管疾患の危険因子は、COVID-19の重症度と死亡率を高める危険因子でもあります。 ここで説明するように、腸内細菌叢は、免疫系、心血管疾患、および肺の健康に関連する応答を調節します。これらのメカニズムは、現在、よりよく理解されています。 これにより、腸内細菌叢の健康をプロバイオティクスでターゲットにする可能性が開かれます。これは、どの機能が主に健康上の利点に関与しているのかが明らかになりつつあるためです。 |
| 2.腸内細菌叢と心血管疾患 |
腸内細菌叢と心血管疾患の間に関係があることは十分に確立されています。 ヒト腸内細菌叢に関する最も詳細な洞察は、メタゲノム調査から得られます。 ディープシーケンシングまたはショットガンメタゲノミクスによって、特定のサンプル内のすべての微生物DNAのシーケンスが決定されます。 メタゲノム(ショットガン)シーケンシングは、正常な(健康な)腸内細菌叢を定義するために使用されてきました[9]。 このような研究は、この方法が細菌群集の配列決定の深さの欠如に悩まされているにもかかわらず、個体間で大きなばらつきがあることを示しており、そのため、存在量の少ない種が見落とされる可能性があります。 ヒトマイクロバイオームは、「ユニバーサル」プライマーを使用したPCRによって増幅された16SrRNA遺伝子フラグメントを配列決定することによってより頻繁に研究されます。 これらの短い16Sフラグメントからのシーケンスは、ショットガンシーケンスよりもはるかに少ない生物情報を明らかにするため、正確な種を特定できないことがよくあります。 |
| 多くの調査により、多くの場合、心血管疾患患者を健康な対照と比較することにより、心血管疾患患者からの便の微生物叢配列が決定されました。利用可能な文献には、患者集団間のかなりのばらつきと転帰が存在しますが、広範な脳卒中では、疾患がない場合、ヒトの腸はバクテロイデス門のメンバーによって支配され、次にファーミキューテスが続き、はるかに少ない数でプロテオバクテリアと放線菌が存在します。細菌の門は何百もの種を表す可能性があり、そのうちのいくつかは人間の腸に見られ、多種多様な「マイクロバイオーム機能」を持っています。種内でさえ(微生物叢を記述する際の精度の範囲の反対側で)、個々の菌株は依然としてそれらの特性がかなり異なる可能性があります。たとえば、Escherichia coliの菌株のゲノムは、数百万の塩基対によってサイズが異なる可能性があり、この種には共生菌株と病原菌株の両方が含まれています。したがって、優勢な門が特定されたとしても、実際の種とその機能的特徴は個体間でかなり異なる可能性があります。 |
| 心血管疾患の症例を健康な対照と比較すると、ほとんどのメタゲノム研究では、ファーミキューテス門とバクテロイデス門の数が増加し、心血管疾患の症例でローズブリア(R. intestinalis)またはフィーカリバクテリウム(F. prausnitzii)の数が減少したことが報告されています。 これらの違いを常に報告するとは限りません[10]。 さらに、ファーミキューテス内では、ラクトバチルス、エンテロコッカス、およびストレプトコッカス種を含むラクトバチルスが、心血管疾患患者においてより豊富であることが多い。 |
| 微生物叢の種の多様性の減少は健康上のリスクと見なされ、種の豊富さの減少は心血管疾患疾患に関連していることがわかりました。 バクテロイデス門の少なくとも一部のメンバーの存在量の減少は、心血管疾患患者、特に肥満、糖尿病、高血圧のリスクのある患者で頻繁に観察されます。 グラム陰性病原菌の存在量が多いことが、心血管疾患患者で観察されることがあります。 |
| 腸内細菌叢が心血管疾患にどのように関与するかについての2つの主要なメカニズムが提案されています。 これらは相互に排他的ではありません。 1つ目はトリメチルアミン-N-オキシド(TMAO)の形成に依存し、2つ目は短鎖脂肪酸(SCFA)の生成に依存します。 |
| 血漿トリメチルアミン-N-オキシドレベルと心血管疾患または血栓性疾患のリスクとの間には明確な相関関係があります[13,14]。 トリメチルアミン-N-オキシドレベルは肥満とも相関しています[14]。 19の研究のメタアナリシスは、トリメチルアミン-N-オキシドレベルの上昇が主要な有害な心血管疾患イベントのリスクの増加と関連していると結論付けました[15]。 腸内細菌叢が心臓の状態にどのように影響するかについてのすべてのメカニズムの説明の中で、トリメチルアミン-N-オキシドの作用は「腸内細菌叢とアテローム性動脈硬化性心臓病との間の最初の潜在的に直接的な関連」として説明されています[16]。 |
| 腸内細菌叢と心血管疾患の間の他のよく認識されているメカニズムのリンクは、細菌が食物繊維を発酵させるときに形成される短鎖脂肪酸の生成です。特に、短鎖脂肪酸/酪酸は腸上皮の完全性を刺激するため、健康な腸に寄与すると考えられています。腸の上皮が損傷すると、細菌のリポ多糖(LPS、エンドトキシンとも呼ばれます)が漏れ出して血流に入り(内毒素血症)、強力な炎症誘発性活性があります。一時的な内毒素血症は、高脂肪の食事の後は正常です。これは、リポ多糖が脂肪と一緒に腸の膜を通過するときに起こります[17]。このようなリポ多糖は、肝臓に輸送する特異的結合タンパク質によって血流からすばやく除去されます。 しかし、慢性的な内毒素血症は、やや不正確に「リーキーガット」と呼ばれるものの結果として、内毒素血症の結果として軽度の炎症の永続的な段階をもたらします。 2型糖尿病や心血管疾患のリスクの増加など、一般的な健康に悪影響を及ぼします[18,19]。 「善玉」腸内細菌によって産生される酪酸は、腸上皮バリアの再生を助け、リポ多糖の漏出を減らします[20]。 |
| 短鎖脂肪酸は主に、宿主が消化できない食物繊維に豊富に含まれる炭水化物を分解するバクテリアによって結腸で生成されます[20]。食物繊維の一般的な健康増進効果は広く受け入れられており、メタアナリシスによって裏付けられています[22]。結腸細菌は、約15%の酪酸を含む短鎖脂肪酸の混合物を生成し[20]、結腸内腔の弱酸性pHをもたらします。酪酸は一般的な腸内細菌によって産生されるため注目を集めており[20,23]、腸内膜の局所的な損傷の修復を刺激することで腸上皮の完全性を刺激します[20,23]。腸の完全性における酪酸産生細菌の役割は、腸内毒素症を一般的に伴う慢性状態である炎症性腸疾患(IBD)が腸内の低短鎖脂肪酸/酪酸産生と関連しており、短鎖脂肪酸補給で改善できるという観察結果とも一致します[ 20]。実際、高繊維、低脂肪の食事から低繊維、高脂肪の食事に切り替えると、血清炎症マーカーが増加し、低繊維の食事は腸内の酪酸レベルを低下させます[24]。多くの一般的な腸内細菌種が酪酸を産生することができ、その中で、Faecalibacterium prausnitziiとEubacterium rectale(両方ともClostridiaメンバー、Firmicutes)が量的に最も重要です。これらのそれぞれは、健康な腸内細菌叢の5〜10%を構成する可能性があります[25]。酪酸を生成する可能性のあるこれらおよび他のファーミキューテスの存在量の違いは、心血管疾患患者と対照の間で示されています。腸内細菌群集は複雑な代謝ネットワークを形成するため、1つの官能基の存在量が他の基の存在量に影響を与える可能性があります。したがって、酪酸産生細菌の存在は、混合培養発酵実験で示されたように、ビフィズス菌などの他の細菌の存在量を操作することによって影響を受ける可能性があります[26、27]。 |
| 心臓と腸の相互作用を複雑にするために、心機能障害は腸機能を損傷し、腸の漏れを引き起こし、内毒素血症を引き起こし、心臓に再び悪影響を与える可能性があります。 腸の虚血は、腸の静脈圧の上昇と血流の減少の結果として発症する可能性があります[28]。 結果として生じる腸の生理機能の変化(ナトリウム分泌や電解質吸収など)は、腸内細菌叢の機能と組成に影響を与える可能性があり、そのようなフィードバックループが適切に配置されていると、原因と結果を解明することは難しい場合がありますが、腸内細菌叢の変化は慢性心不全を伴うことがよくあります。 |
| 3.腸内細菌叢と肺の健康 |
胚発生の間、腸と肺の両方が同じ原始的な前腸から形成されます。 出生後、両方の器官は、より深い組織と外部環境との間の粘膜バリアとして機能し、両方とも局所的な微生物集団を抑制しなければなりません。 同時に、それらは必須栄養素(腸)とガス(肺)の通過を可能にしなければなりません[29]。 発育中の胎児はすでに微生物叢にさらされている可能性がありますが、これについては議論されています[30]。 とにかく、出生直後、肺と腸の両方に初期の微生物叢が定着しており、優勢な細菌門に関しては、両方の器官でその段階で互いに類似しています[29,31]。 |
| 特定の年齢以降、健康な状態では、肺には、腸内細菌叢と同様に、細菌門バクテロイデス門とファーミキューテス門が優勢な特定の微生物叢がありますが、優勢な属と種はこれらの臓器間で異なる場合があります。しかし、腸には、肺よりも組織1グラムあたり少なくとも100万個多くの細菌細胞が含まれています[29]。これらの臓器は通常の微生物叢を感知して通信し、この宿主と微生物の相互作用の背後にあるメカニズム、腸と肺が病原体にどのように反応するか、そしてそれらが一時的な損傷をどのように回復するかはすべて驚くほど似ています。腸粘膜と気道粘膜の両方が循環リンパ球によってパトロールされます。リンパ球は、これらの細胞が置かれたままではないため、2つの臓器間の直接的な免疫学的接続として機能します。 特定の抗原への曝露は、局所的な樹状細胞によって感知されます。 これは、Tヘルパー細胞1型(Th1)対Th2応答などの将来の免疫応答を形成し、それぞれの微生物叢が重要な役割を果たす健康と病気に長年の影響を与えます。 これらの臓器が「通信」するクロストークは、「腸-肺軸」と呼ばれます。 |
| 腸と肺の間の臓器間コミュニケーションのほとんどは、血流を介して行われます。臓器間効果を発揮する免疫応答には、病原体や毒素の局所的な存在に応答する抗体活性化Tリンパ球、および炎症反応を必要なレベルのみに制限するために産生される抗炎症性サイトカイン(糖質コルチコイド)が含まれます[32 ]。糖質コルチコイド(GC)は一般に抗炎症機能を持っています(抗炎症薬デキサメタゾンは合成糖質コルチコイドです)。糖質コルチコイドは副腎によって生成されるだけでなく、腸と肺の上皮によって少量生成され、トリガー後の過剰反応を回避するために局所免疫細胞応答をダウンレギュレーションします。炎症誘発性腫瘍壊死因子(TNF)が腸で産生されると、同時に糖質コルチコイドの産生を刺激し、バランスの取れた炎症反応を保証する「ブレーキ」を開始します[32]。免疫学的防御と抗炎症性の「ブレーキ」の両方が、局所的に感染した領域を超えて到達し、腸と肺の間のクロストークを可能にします。さらに、腸で生成された短鎖脂肪酸は血流を介して肺に到達し、腸で行われるのと同じように肺細胞の健康を改善することができます。 短鎖脂肪酸は、さまざまな細胞型で発現するGタンパク質共役型受容体(GPVCR)に結合します。 Gタンパク質共役型受容体はシグナル伝達システムとも結合しており、組織や細胞の種類に応じて、短鎖脂肪酸が複数の効果を発揮する方法を説明しています。これは他の場所でより深く扱われます[33]。 |
| 明らかに、腸と肺の両方が病原体に感染する可能性があり、一方の感染が他方の健康に影響を与える可能性があります。 腸管に存在する微生物群集は、呼吸器疾患の結果として変化する可能性があり、同様に、腸内細菌叢の変化は、感染に対する肺の感受性に影響を与える可能性があります。 ウイルス性および細菌性呼吸器感染症は、腸内毒素症および腸症状を引き起こす可能性があります[34]。 反対方向への影響の例として、気道感染症の発生率または重症度は腸内細菌叢の影響を受けます[35]。 これは、図1の点線の矢印で示されています。 |
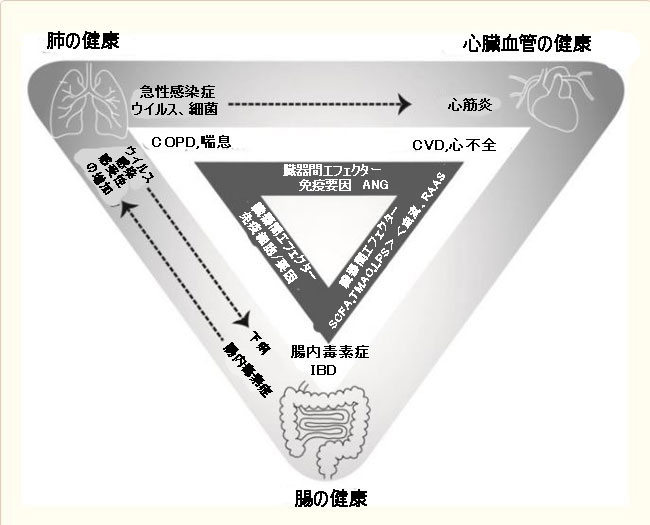 |
| 図1 肺の健康、心臓血管の健康、腸の健康の相互作用 |
点線の矢印は、局所急性感染症の臓器間効果を示しています。 白い三角形は主な慢性状態を示し、暗い三角形はコミュニケーションを可能にする主要なエフェクターのいくつかを示しています。 略語:COPD:慢性閉塞性肺疾患、CVD:心血管疾患、IBD:潰瘍性大腸炎、 ANG:アンジオテンシンSCFA:短鎖脂肪酸、 TMAO:トリメチルアミン-N-オキシド、LPS:リポ多糖、、 RAAS:レニン・アンジオテンシン・アルドステロンシステム |
| 上記のように、病原体の存在は、どちらかの臓器の免疫細胞によって認識されます。局所的には、この認識は主にパターン認識受容体(PPR)に結合する病原体関連分子パターン(PAMP)の結果です。 Toll様受容体(TLR)などのこれらのパターン認識受容体は、病原体が検出された場合に自然免疫応答を開始する主要な「スカウト」として機能します。免疫系のこの強力な枝は、RNAやリポ多糖などの高分子の保存されたモチーフを認識しますが、壊死した宿主細胞によって放出されるシグナルにも応答します。結局のところ、感染は細胞の損傷を伴い、病原体がない場合でも、損傷した細胞は有害である可能性があり、排除する必要があります。腸内のToll様受容体は、病原体と共生生物の存在に同様に気づきますが、病原体関連分子パターンまたはその他の危険関連分子パターン(DAMP)と無害な微生物MAMPを区別することができます。このように病原体が認識されると、適応免疫応答が活性化され、粘膜免疫に関与するIgAを含む病原体特異的抗体の産生が開始されます。 |
| 腸内細菌叢が抗生物質によって変化するマウスで、腸と肺の間の相互接続が詳細に調査されています。 この治療により、インフルエンザウイルスなどの呼吸器感染症に対する感受性が高まります。 さらに、腸内細菌叢を人工的に欠いている非生物的マウスは、肺胞にマクロファージを持ち、転写産物が変化し、細菌を貪食する能力が低くなります。これは、腸内細菌叢が肺の正しいマクロファージ機能に不可欠であることを示しています。 肺と腸の間の相互作用は、年齢層の両端で特に重要です。たとえば、農場で育った結果としての幼い頃の多様な腸内細菌叢は、アレルギー性喘息を予防します[36]。 高齢化社会における腸内毒素症の一般的な状態(食事の多様性の制限に関連する)は、肺感染症に対する感受性の増加と相関することがわかっています[37]。 |
| 4. 肺と心臓のつながり |
多くの急性ウイルス性および細菌性肺感染症は心筋炎を引き起こす可能性があり、ウイルス性気道感染症はこの状態のより一般的な原因の1つです。 一部のウイルスは心臓に感染する可能性があり(一部のヘルペスウイルスまたはエンテロウイルス種のように)、他のウイルス(インフルエンザウイルスなど)は過剰反応性免疫応答の結果として心筋炎を誘発します[38]。 彼らのレビューでは、Favere et alは、ウイルス性心筋炎の発症におけるToll様受容体の役割について詳細に論じています。 心筋炎を引き起こすウイルス種のほとんどは非常に一般的ですが、動物実験は、遺伝的素因がこの潜在的に深刻な合併症の引き金となる可能性があることを示唆しています。 |
| 別のメカニズムは、肺のSARS-CoV-2感染とそれに伴う心臓合併症との関連の背後にあります。 SARS-CoV-2は主に上気道に感染し、細胞侵入の主なドッキング受容体はアンジオテンシン変換酵素2(ACE2)です(他のタンパク質も必要ですが、ここでは簡単にするために無視します)。 アンジオテンシン変換酵素2は多くの組織で発現する膜貫通タンパク質です。それは、一連の重要な経路を調節するペプチドに対して重要なエキソペプチダーゼ酵素活性を持っています。これは、血圧と体液の恒常性を調節するレニン-アンジオテンシンシステムの重要な要素としてよく知られています。 アンジオテンシン変換酵素2の主な基質は、8アミノ酸長のペプチドであるアンジオテンシンII(Ang II)であり、アンジオテンシン変換酵素2は7 merのアンジオテンシン(1-7)に短縮されます(図2、パネルA)。 アンジオテンシンIIは血管収縮剤として機能しますが、アンジオテンシン変換酵素2製品のアンジオテンシン(1-7)は血管拡張剤です。その前駆体であるアンジオテンシノーゲンは、肝臓でプレプロタンパク質として産生され、腎レニンの活性によってデカペプチド(アンジオテンシンI)として放出され、その後、ペプチダーゼアンジオテンシン変換酵素1(従来は単にアンジオテンシン変換酵素(ACE)と呼ばれていました)によって活性オクタマーアンジオテンシンIIに切断されます。 八量体のアンジオテンシンIIは血流を循環し、必要に応じてアンジオテンシン変換酵素2の基質として機能します。これは、血管上皮を含む多くの組織によって発現されます(「脱落」の結果として、少量の機能的に活性なアンジオテンシン変換酵素2も循環しています。さらに別のタイプのプロテアーゼによって実行される)。 |
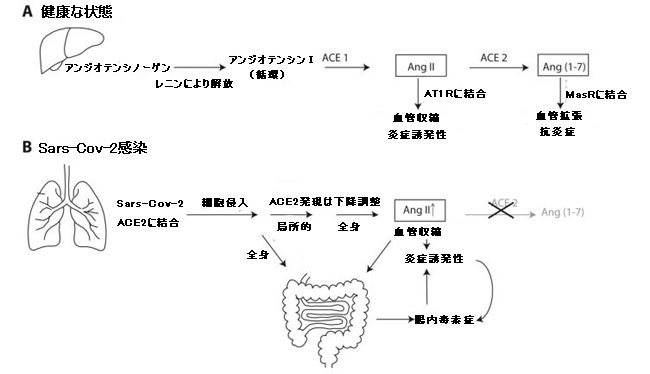 |
| 図2 健康およびSARS-CoV-2感染時のレニン-アンジオテンシンシステム(RAS)の一部としてのアンジオテンシン変換酵素1(ACE1)およびアンジオテンシン変換酵素2(ACE2)の役割 |
| パネルAは、肝臓で生成された後のアンジオテンシノーゲンからのアンジオテンシン IIおよびアンジオテンシン(1-7)の生成を示しています。 血圧と炎症に対するそれらの拮抗作用が示されています。 アンジオテンシン1からアンジオテンシンIIを与える代替経路は示されておらず、簡単にするために代替成分、受容体、および経路は省略されています。 パネルBは、SARS-CoV-2感染時のアンジオテンシン変換酵素2抑制の炎症誘発性効果をまとめたものです。 腸は正のフィードバックループで炎症誘発性状態を増加させます。 |
| アンジオテンシン変換酵素2がアンジオテンシンIIをアンジオテンシン(1-7)に変換した後、このペプチドは細動脈平滑筋細胞のMas受容体(MasR)に結合できるようになり、血管弛緩と血圧の低下をもたらします[39]。 したがって、広い脳卒中では、アンジオテンシン変換酵素2活性が高いと血圧が下がり、アンジオテンシン変換酵素2活性が低いと血圧が上がります。 アンジオテンシン変換酵素1とアンジオテンシン変換酵素2は、2つの中和ホルモンであるアンジオテンシン IIとアンジオテンシン(1-7)のバランスをとっています(図1)。 実際には、明確にするためにここでは省略されている、はるかに多くの酵素と受容体が関与しています。 読者は、より詳細な情報について他のレビューに導かれます[39,40]。 |
| アンジオテンシンIIとアンジオテンシン(1-7)の同様の対抗作用は、炎症に対するそれらの効果について観察されます。一般的に言えば、アンジオテンシン IIは炎症誘発性であり、アンジオテンシン(1-7)は抗炎症性です(ここでも、ここで説明できるよりもはるかに多くの要因と影響が関係しています)。 アンジオテンシン変換酵素1酵素とアンジオテンシン変換酵素2酵素はどちらも、細胞の成長と増殖、炎症、およびサイトカイン産生の局所的(臓器特異的)調節に関与しており、ほとんど反対の効果があります[41]。 アンジオテンシン変換酵素2はまた、受容体特性(膜貫通および細胞質尾部相互作用を介して)、ならびにトランスポーターとのペアリングによる膜貫通アミノ酸輸送などの重要な機能におけるヘテロダイマーパートナーとしての調節機能を持っています。 このように、それは炎症性腸疾患[42]で役割を果たします。 |
| アンジオテンシン変換酵素2は、ヒトの上気道および下気道の特定のタイプの上皮細胞の頂端表面に高レベルで発現し、一般的で病原性の低いコロナウイルスを含む呼吸器ウイルスの受容体として機能します。これは、高病原性SARS(現在はSARS-CoV-1と呼ばれています)およびMERSで最初に実証されました[43,44]。 COVID-19の原因であるSARS-CoV-2は、そのスパイクタンパク質によって1対1の分子ベースでアンジオテンシン変換酵素2に熱心に結合します。肺上皮の頂端表面で利用可能なアンジオテンシン変換酵素2分子の量は、それらに到達するウイルス粒子の数をはるかに超えているため、最初は、比較的少数のウイルス粒子の結合が利用可能なアンジオテンシン変換酵素2分子の数に直接影響することはありません。 しかし、SARS-CoV-2が細胞(追加の酵素が必要)に入るとすぐに、アンジオテンシン変換酵素2の発現は、そのde novo産生、膜貫通挿入、および細胞表面への輸送の観点から、特定のウイルスタンパク質に対する細胞応答によってダウンレギュレーションされます。 これは、感染した細胞だけでなく、抗ウイルス性のパラクリンシグナル伝達を介して周囲の細胞でも起こります。これは、局所組織が、追加の入ってくるウイルス粒子のドッキングポイントの数を制限し、感染した細胞から逃げるウイルスの最終的な局所拡散を制限しようとする方法です。同時に、血管弛緩および抗炎症性アンジオテンシン変換酵素2機能の量が減少するため、アンジオテンシン変換酵素1機能不全はその血管収縮および炎症誘発作用を発揮することができます。 このようにして、ウイルスによる損傷は感染部位に隔離され、順次局所的、次に領域的な自然免疫応答経路が開始されます(図2、パネルB)。 |
| 気道でのアンジオテンシン変換酵素2の発現には多くの注意が払われていますが、腸系と胆嚢では対数桁の高レベルで発現しています[45]。 少なくともマウスでは、アンジオテンシン変換酵素2の欠乏が炎症を引き起こし、腸内細菌叢に有害な変化をもたらすことが実証されています[46]。これが、COVID-19の軽度の症例でさえ胃腸症状を伴うことが多い理由かもしれません。 腸内では、アンジオテンシン変換酵素2エキソペプチダーゼ活性が腸上皮および神経内分泌細胞による炎症誘発性ペプチドの局所産生を制御します。 さらに、免疫細胞による抗菌ペプチドの放出を可能にし、それらが組み合わさってマイクロバイオームの毎日の健康的な組成に影響を与えます[47]。 アンジオテンシン変換酵素2はまた、アミノ酸トランスポーターによる腸のトリプトファン取り込みの調節を変えることによって免疫応答を調節します。 要約すると、アンジオテンシン変換酵素2は、レニン-アンジオテンシンシステム(RAS)の重要な要素であることに加えて、アミノ酸の恒常性、自然免疫、および腸(そしておそらく肺)の微生物生態学の重要な調節因子です。 |
| 5.病状から学ぶ |
| 心血管疾患に関する前のセクションは、病状の研究が健康に関する洞察にもつながる可能性があることを示しています。 これは、腸と肺の両方の慢性疾患状態に当てはまります。 クローン病や潰瘍性大腸炎などの慢性炎症性腸疾患(IBD)、および肺の慢性閉塞性肺疾患(COPD)は、持続性の局所腸内毒素症と関連しており、同様の根本的なメカニズムとリスク要因があります。 両方の慢性障害は、喫煙と特定の遺伝的素因に関連しています。 |
| 慢性炎症性腸疾患のような腸の慢性疾患状態は、ほとんどの場合、長期の微生物叢の腸内毒素症を伴い、慢性閉塞性肺疾患の有病率が増加し、その逆もあり[34]、湿疹との関連も明らかです。 前に述べたように、慢性炎症性腸疾患は短鎖脂肪酸補給によって改善することができます。 マウスの実験で実証されたように、食物繊維と短鎖脂肪酸の両方の経口摂取は、気道の恒常性に寄与する可能性があります[32]。 この文脈では、慢性炎症性腸疾患と心血管疾患の間に関連性も存在するのは当然のことです。 台湾でのコホート研究は、重度の慢性炎症性腸疾患患者における末梢動脈疾患(PAD)のリスクが27.5倍高いことを示しました[48]。 |
| 腸内細菌叢の組成の破壊(内的または外的要因による)は、免疫系の自己制限能力を低下させる可能性があり、これは腸自体だけでなく、慢性閉塞性肺疾患および喘息を引き起こすことによって慢性炎症状態を引き起こす可能性があります[33]。 確かに、喘息患者と対照の間の腸内微生物叢の組成の違いが実証されています[49]。 腸内細菌叢の組成を破壊する可能性のある外的要因には、食事、特定の毒素と病原体、および投薬が含まれます。 これらのトリガーのいずれかが腸内毒素症を引き起こす可能性があり、局所的および全身的な炎症につながる腸免疫の望ましくない影響をもたらす可能性があります。 |
| 腸内毒素症も肥満と糖尿病に関連しており、肥満の人は炎症誘発性状態が高く、喘息の発生率が高くなりますが、体重減少(たとえば減量手術による)は喘息を減らすことができます[50,51]。 肺上皮は、IgA抗体、ディフェンシン、およびリゾチームの局所免疫活性を制御し、Th2型炎症を刺激するサイトカインを介してそれらの産生を調節します。これは、喘息の発症に関係しています[49]。 腸内細菌叢はTh2免疫応答に影響を与えるため、喘息の発症に影響を与える可能性がありますが、この状態は主に吸入に起因すると考えられています。 |
| 6.理想的なプロバイオティクスにはどのような活動が必要ですか? |
| これまで、さまざまな健康状態または病状と腸内微生物叢の観察との相関関係が言及されており、臓器間クロストークのメカニズムが簡単に概説されています。 まだ答えられていないより大きな問題は、腸内細菌叢を操作して、個人の健康状態を改善できるかどうかということです。 これはプロバイオティクスが行うと考えられていることです。 |
| この文脈で最初に気付くのは、体が非病原性の腸内細菌叢とプロバイオティクス細菌を区別しないということです。腸内の共生菌または一時的に通過する細菌種の存在は、上記の宿主と微生物の相互作用を介して「通知」されますが、害がない限り(病原体関連分子パターンを発現し、警報を発する局所的な細胞損傷を引き起こす)、これらの細菌はそれらが他の種に取って代わられるまで、栄養素と生きるためのスペースを求めて互いに競争するために、事実上放っておかれます。次に注意すべきことは、プロバイオティクス能力について調査されたほとんどの細菌種は、腸の正常な住民であるだけでなく、通常、食品発酵にも使用されているということです。これらの選択は部分的に歴史的であり(最初のプロバイオティクス活動は低温殺菌されていないヨーグルトに存在する細菌に起因していました)、部分的に実用的です(スターターカルチャーは安全であると考えられており、プロバイオティクスとして適用するのが簡単です)が、プロバイオティクス活性の調査に最適な種を選択しました。実際、過去10年間で多くのレビューペーパーが安全性の絶対的な要件について論じてきましたが、おそらくその目標を達成するのが非常に難しいため、機能性に取り組んだり評価したりしたものははるかに少ないです。文献で多く議論されている他の要件は、プロバイオティクス細菌が生きたまま腸に到達し、そこに住み、そこで増殖する必要があるということです。 つまり、(主にプロバイオティクスの一般的に受け入れられている定義の一部であるため)未定義の期間「コロニー化」しますが、長期的なコロニー形成が望ましいか回避すべきかについては議論が続いています。 彼らの提案された行動に照らして、死んだバクテリアがどのように健康に持続的なプラスの効果をもたらすことができるか想像するのは難しいです、それにもかかわらず過去に提案されました[52]。 最後に、プロバイオティクス細菌は通常、単一培養として工業的に生産されます。これにより、品質管理と培養の安定性が保証されますが、一部の市販製品には菌株または種の混合物が含まれています。しかし、腸のより重要な有益な細菌種のいくつかは、主要な酪酸生産者のいくつかを含めて、単一栽培で繁殖するのが困難です[25]。しかし、これらは混合培養で増殖する可能性があり、その結果、酪酸の生産が増加します[53,54]。規制上の制限により、混合培養生産の使用が制限されていますが、プロバイオティクス株と種の組み合わせカクテルの有益な可能性が明らかであるため、そのような規制を検討する時期かもしれません。 |
| 通常の乳酸菌の代わりに、プロバイオティクスの使用のために健康な人の腸ですでに一般的に見られる細菌種を選択する必要がありますか?細菌種が人間の腸で一般的に見られるとしても、すべての個体がその種によって等しいレベルにコロニーを形成するという保証はなく、同じように利益を得るという保証もありません。大腸菌もまた重要な例です。腸内に大量に見られることも、すべての個体が大腸菌によって容易にコロニーを形成することもありません[55]。健康な人に関連しているが、肥満、糖尿病、心臓代謝性疾患、および軽度の炎症を患っている人には見当たらない、または非常に少ない量で存在する、あまり馴染みのないグラム陰性菌種の別の例は、腸粘膜層にコロニーを形成するAkkermansia muciniphilaです。そして、プロバイオティクス候補として提案されています[56]。あまり認識されていないグラム陽性菌の例として、 Enterococcus faecalis株は子供の副鼻腔炎を予防するのに効果的であることが示されています[57]。さらに、ここで説明する例が示すように、腸内毒素症に関連するさまざまな病状がありますが、これらすべてが、影響を受けるすべての個人の1種類の細菌プロバイオティクスで改善できると仮定するのは単純です。 |
| 個人の微生物叢の変動は広範囲であり、時間とともに変化し、一人の人の上腸と下腸の間でさえも変化します。これは、広く活性なプロバイオティクス細菌株の知識に基づいた推測をさらに困難にします。ただし、ここでは、基礎を提供する可能性のあるいくつかの一般的な観察結果について繰り返し言及しています。短鎖脂肪酸を生成する種、特に酪酸は、複数の点で有益であるようです。 短鎖脂肪酸の有益な活性は、すでに述べたものを超えており、腸のIgA産生の上昇、T-reg細胞分化の活性化、免疫細胞のエピジェネティックな変化の調節も含まれます[58]。さらに、抗炎症プロセスを活性化し、炎症誘発作用を制限する細菌は、病気のアトピー状態の免疫応答を和らげるのに役立ちます。 最後に、トリメチルアミン-N-オキシドレダクターゼを欠く細菌、またはこの酵素を発現する他の細菌の数を減らす細菌は、トリメチルアミン-N-オキシドの生成を制限し、これは非常に有益であるはずです。図3に要約されているこれらの機能は、プロバイオティクス細菌の健康上の利点を説明するために頻繁に提示されるメカニズムの説明とは部分的に異なります。それらの伝統的に考えられているプロバイオティクス機能には、病原体との競争と病原体の除去が含まれます。このような拮抗作用は、Alfred NissleがE.coli Nissle1917を分離した最初の基準でした[55]。他のモードには、病原体が粘膜表面に付着するのを阻害すること、乳糖の消費と乳酸の生成、および抗酸化活性が含まれます[59,60]。 |
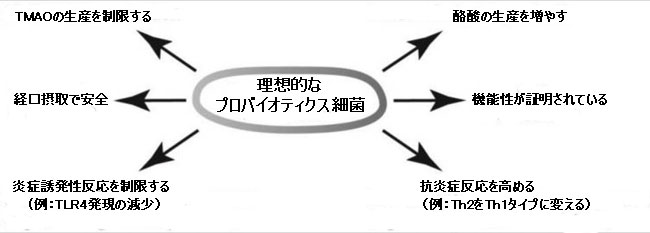 |
| 図3 理想的なプロバイオティクス細菌の必要な活動は、それらが腸を超えた臓器の健康に利益をもたらすことができるように、メカニズムの理解に基づいて提案することができます。 |
| 酪酸生産の重要性は長い間確立されてきましたが[61]、腸の障害に焦点を当てることで、その潜在能力の評価が制限された可能性があります。プロバイオティクスが呼吸器感染症の予防に役立つことは現在十分に確立されており、これはメタアナリシスで確認されています[62,63]。免疫調節のメカニズムは、現在よりよく理解されています[64]。「免疫系の刺激」などの一般化の代わりに、全身効果をより正確に説明できるようになりました。 シーンは現在、腸にコロニーを形成する可能性のある細菌でそのような活動(図3に要約されている)を検索するように設定されており、それによって既知のプロバイオティクスの原型をはるかに超えて調査を拡張します。新しいプロバイオティクス候補の検索は、以前に述べられたように、理想的には、薬物やワクチンに必要なものと同様に、ランダム化臨床試験に基づいている必要があります[52]。 EUを含む世界の一部の地域では、これらの要件は、健康上の利点を主張する新規製品を登録するためにすでに実施されています。 |
| しかし、既知の安全なバクテリアをテストして、それらが持つ可能性のある有益な効果をテストする代わりに、臨床試験は費用がかかるため、機能性と組み合わせた安全性の評価(適切な用量とともに、臨床試験で本質的に対処される2つの目標)によってプロバイオティクスの開発に向けた取り組みがほとんど開始されていない理由である可能性があります。 機能性を証明する必要がある場合、プロバイオティクス細菌が生きたまま腸に到達してそこでコロニーを形成する必要があるかどうかの議論は、コロニー形成する能力がなければ、そのような証明を提供するのは難しいため、最終的に明確な結論に達する可能性があります。 |
| プロバイオティクスの通常の容疑者を対象に実施されたランダム化臨床試験の例はより豊富であり、ここで関連性の条件に対するそれらの影響を評価した多数のメタアナリシスが公開されています。これらは表1にまとめられています。メタアナリシスは過去の研究の結果を累積的に組み合わせているため、2016年以降に公開されたもののみが表に含まれています。全体的な有意な効果が見られなかったものと比較して、さまざまな条件で、はるかに多くのメタアナリシスがプロバイオティクスの効果を見つけました。報告された可能性のあるサブグループ分析は、ここでは考慮されません。この種の研究には注意すべき制限があります。メタアナリシスで得点される効果はしばしば軽微であり、研究デザインと調査された試験の質はかなり異なり、試験はしばしば非常に不均一であり、複数のプロバイオティクス種、株、用量がテストされます。それにもかかわらず、ここで関心のある条件に対するプロバイオティクスの機能を報告する圧倒的な数の臨床試験が現在公開されていると結論付けます。 |
 |
| 7.プロバイオティクスとCOVID-19 |
| プロバイオティクスがSARS-CoV-2感染またはCOVID-19症状を完全に防ぐことができると考えるのは素朴ですが、健康な腸内細菌は複数の臓器に明らかな健康上の利点があるため、プロバイオティクスはSARS-CoV-2感染を改善するための複数の戦略の1つとして提案されています [6,114,115,116,117,118]。 |
| 中国からの初期の観察は、一部のCOVID-19患者が、腸内のラクトコッカス属とビフィズス菌属の種が少ない腸内毒素症に苦しんでいることを示唆しています。 SARS-CoV-2に感染すると、肺だけでなく腸内でもアンジオテンシン変換酵素2の発現がダウンレギュレートされます[45]。アンジオテンシン変換酵素2は腸内での抗菌ペプチドの局所産生を厳密に制御するため、微生物叢に直接影響を与える可能性があります。 腸内毒素症について提唱されている別の説明は、腸上皮組織におけるウイルス複製です。 ただし、長期のウイルスゲノムRNA排出は便で検出されることがありますが、腸内での活発な複製は大規模には行われません。 |
| 中国からの初期の観察は、一部のCOVID-19患者が、腸内のラクトコッカス属とビフィズス菌属の種が少ない腸内毒素症に苦しんでいることを示唆しています。 SARS-CoV-2に感染すると、肺だけでなく腸内でもアンジオテンシン変換酵素2の発現がダウンレギュレートされます[45]。アンジオテンシン変換酵素2は腸内での抗菌ペプチドの局所産生を厳密に制御するため、微生物叢に直接影響を与える可能性があります。 腸内毒素症について提唱されている別の説明は、腸上皮組織におけるウイルス複製です。 ただし、長期のウイルスゲノムRNA排出は便で検出されることがありますが、腸内での活発な複製は大規模には行われません。 |
| このような観察結果は、腸に十分なレベルと多様性の免疫調節プロバイオティクス細菌が含まれている場合、個人がCOVID-19症状の発生と重症度を軽減する可能性があることを示唆しています。抗炎症作用のあるプロバイオティクスは、サイトカインストームのリスクを大幅に減らす可能性があります[120]が、急性呼吸窮迫症候群の原因となる一連の炎症性メディエーターもプロバイオティクスによって減らすことができます[116]。図2に示すように、アンジオテンシン変換酵素1の発現を低下させるプロバイオティクスがいくつか報告されています[121]。これはレニン-アンジオテンシンシステムを介した抗炎症反応に寄与します。 |
| さらに、特定の細菌種が腸内のアンジオテンシン変換酵素2発現を調節し、レニン-アンジオテンシンシステムを介して肺に有益な下流効果をもたらす可能性があることが示唆されています[58,122]。 酪酸の生成は、明らかに腸と肺の上皮内層の損傷を修復するのに役立ちます。全身性の抗ウイルス活性を有するI型インターフェロンの定常状態レベルを増加させるいくつかのプロバイオティクスの能力も貢献する可能性があります[65]。ワクチン接種の結果としてパンデミックが減少しているとしても、ワクチンの成功はプロバイオティクスによって強化される可能性があるため、プロバイオティクスがこれを支援する可能性があります[58]。 プロバイオティクスの有益な活性のほとんどは抗炎症性であるように思われるため、これは直感に反しているようです。 しかし、健康でバランスの取れた免疫システムは、ワクチンに対してより完全かつ正確に反応し、ワクチン接種に対するプロバイオティクスのプラスの効果は、リポ多糖のアジュバントのような効果に部分的に起因している可能性があります[123]。 |
| 最後に、SARS-CoV-2感染の急性期に比較的軽度の症状しか経験しなかった、または症状がなかった患者は、後に「長いCOVID-19」と総称されるさまざまな症状を長期間発症する可能性があります[124 ]。 この感染後の続編の本当の負担はまだ明らかではありません。 症状は数ヶ月続く可能性がありますが、事例証拠は、COVID-19ワクチン接種を受けた後に状態が改善する可能性があることを示唆しています。 少なくとも一部の個人では、複雑な免疫システムの機能不全のチェックとバランスは、おそらく免疫調節を介して、正しい入力によってリセットされる可能性があるようです。 この文脈では、プロバイオティクスが長いCOVID-19患者のより速い回復を助けることができるかどうかを見ることは興味深いでしょう。 |
| 8.結論 |
| 何十年もの間、プロバイオティクスはさまざまな病状の解決策として提案されてきました。それにより、重度の慢性疾患から一般的で未定義の「健康の改善」に至るまで、変化し続ける問題の解決策を提供します。これは当然のことながら、特に腸以外の臓器での活動が標的にされたと思われる場合には、ある程度の懐疑論をもたらしました。しかし、ケモカイン、サイトカイン、神経内分泌分子、移動する免疫細胞、特定の代謝物、さらには神経系を介した臓器の相互接続性をよりよく理解することで、腸内の細菌が実際に広範囲に及ぶ可能性があることが明らかになりつつあります。全身的な結果、そして健康な腸内細菌叢は、多くの状態の予防、緩和、および治癒に貢献することができます。斬新な洞察はまた、理想的なプロバイオティクスのためのより明確な「ウィッシュリスト」をもたらしました。通常の候補はそれらの機能特性以外の基準に基づいて選択されたため、さまざまな条件で最も有益な細菌の最良の候補をまだ特定している可能性は低いです。当然のことながら、安全性が最も重要な基準であり、経口摂取時にすでに安全であると考えられていた工業的に使用される細菌が論理的な第一選択の候補でした。これらのプロバイオティクス細菌を使って行われた研究は、確かに貴重な洞察につながりました。プロバイオティクス細菌がどのように機能するかを正確に理解し始めた今、次の10年間の研究は、より適切でより強力な候補となる可能性があります。少なくとも、腸内細菌が肺の状態や心臓病に有益であるかどうか、そしてどのように有益であるかは、もはや謎ではありません。これらの新しい提供された洞察を使用して、仮説主導の研究の次の段階を導く場合、より多くのブレークスルーが期待できます。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
| この文献は、Int. J. Mol. Sci. 2021, 22, 9700に掲載されたInteractions between the Gut Microbiome, Lung Conditions,and Coronary Heart Disease and How Probiotics Affect These.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |