ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) | |
ウイルス感染症、マイクロバイオーム、およびプロバイオティクス |
|
Ashton Harper et al., |
|
|
|
|
概要 |
|
| ウイルス感染は、世界中でかなりの罹患率と死亡率を引き起こし続けています。これらの感染症の最近の増加は、気候変動、人や物の移動性の向上、人口動態の急速な変化など、複雑で多因子的な外部要因が原因である可能性があります。これらの外的要因と並行して、ウイルス免疫に関連する内的要因についての理解が深まっています。胃腸(GI)マイクロバイオームは、免疫および宿主防御機構の重要な調節因子として機能し、宿主免疫系の重要なプレーヤーであることがますます示されています。証拠の増加は、胃腸マイクロバイオームと宿主免疫系の間の恒常性の崩壊がウイルス免疫に悪影響を与える可能性があることを示しています。このレビューは、宿主とマイクロバイオームの相互作用が、初期の生命因子、抗生物質への曝露、免疫老化、食事療法、炎症性疾患など、免疫系をどのように形成するかについての理解に光を当てることを目的としています。また、宿主共生生物およびマイクロバイオーム治療薬が、ウイルス性胃腸炎、ウイルス性肝炎、ヒト免疫不全ウイルス(HIV)、ヒトパピローマウイルス(HPV)、ウイルス性上部、呼吸器感染症(URTI)、インフルエンザおよび重症急性呼吸器症候群コロナウイルス2(SARSCov-2)などのウイルス感染の予防および/または治療にどのように影響するかについての証拠ベースについても説明します。胃腸のマイクロバイオーム、侵襲性ウイルス、および宿主の生理機能の間の相互作用は複雑であり、まだ完全に特徴付けられていませんが、マイクロバイオームがウイルス性疾患の結果に影響を与える可能性があることを示す証拠が増えています。現在のエビデンスベースは有益ですが、この複雑な関係の根底にある一連の免疫学的メカニズムを完全に理解するには、さらに適切に設計されたヒトの臨床試験が必要になります。 | |
序章 |
|
|
ウイルス(ラテン語の「ウイルス」毒から)は、すべての進化的実体の中で最も遍在的で豊富です(Norrby.,2008)。 地球上に固有のウイルスがいくつ存在するかはわかりませんが、哺乳類(〜5,500種)だけに感染すると推定されるウイルスは約32万種類(Anthony,2013)、人間に感染するウイルスは1,000種類を超えることが知られています(Lassoetal,2019)。 ウイルスの起源については議論の余地がありますが、ウイルスが最後のユニバーサルセルラー祖先(約40億年前)よりも前に存在し、地球上の生命の進化に重要な役割を果たした可能性があると考えるのは魅力的です。 特に私たち自身(ヒトゲノムの少なくとも8%は感染性レトロウイルスに関連する配列で構成されています)(Moelling and Broecker,2019)。 |
|
| ウイルス感染は、次のようなさまざまな組織や臓器を攻撃します。 上気道および肺(例、ライノウイルスおよびインフルエンザ)、結腸(例、ロタウイルス)、肝臓(例、B型肝炎ウイルス)、脊髄(例、ポリオウイルス)、血管内皮細胞(例、エボラ)、 および白血球(例えば、ヒト免疫不全ウイルス)。 ウイルス感染は、人類の歴史の中で驚異的な数の死の原因となっています。 1980年に根絶に成功した天然痘(天然痘ウイルスによって引き起こされる)は、20世紀だけで3億から5億人の死者を出したと推定されています(Thèvesetal,2014)。 ヒト免疫不全ウイルス(HIV)はこれまでに7500万人に感染し、そのうち3200万人を殺しました。 そして、2020年12月1日の時点で、重症急性呼吸器症候群コロナウイルス2(SARS CoV-2)は、6,300万人以上に感染し、140万人以上が死亡しましたが、症例は急速に増加しています。 この病気の影響はまた、世界経済に深刻な打撃を与えます。 | |
| すべての後生動物(多細胞、ミトコンドリア真核生物)と共通して、私たちは微生物の関連するコミュニティを持つ宿主、すなわちホロビオント(訳者注:宿主とその周辺に生息する他の多くの種の集合体)です(Simon et al.,2019)。定義された環境に存在する微生物(細菌、古細菌、下等および高等真核生物、およびウイルス)の集合体は、マイクロバイオームという用語を定義します。 マイクロバイオームとは、特定のマイクロバイオータ(訳者注:細菌、微生物の集団を意味し、「細菌叢、微生物叢」に近い。以下マイクロバイオータを細菌叢と表記し、マイクロバイオームと区別します)とその集合ゲノムの生息地全体と周囲の環境条件を指します(Marchesi and Ravel,2015)。人間の胃腸(GI)管だけでも、4×1013の細菌が含まれています。これは、私たち自身の細胞の総数よりもわずかに多い数です(Sender et al.,2016)。ホロゲノーム(ホロビオントゲノム)という用語は、宿主ゲノム(ヒト〜20,000遺伝子)と関連する集合微生物ゲノム(> 3,300万遺伝子)の組み合わせを表します(Simon et al.,2019)。そのうち900万個はユニークなタンパク質コード遺伝子です(Yang et al.,2009)。したがって、私たちの微生物の居住者は、私たちの遺伝的レパートリーを大幅に拡大し、私たちだけでは不可能な方法で私たちの環境をナビゲートすることを可能にします。 | |
| 国立衛生研究所によって2007年に開始されたヒトマイクロバイオームプロジェクト(HMP)は、人間の健康と病気におけるマイクロバイオームの重要性の特徴づけと理解を可能にしました(Turnbaugh et al.,2007)。さらに、真核生物の二本鎖DNA(dsDNA)ウイルスは、ヒトマイクロバイオームプロジェクトによって生成されたメタゲノム配列データを調べることによって分析されています。この研究では、各個人で平均5.5のウイルス属が検出され、高い対人多様性が報告されました(Wylie et al.,2014)。人間のウイルス叢に関する研究は、次のような課題すなわちウイルスの一般的なマーカーの欠如、ウイルス叢要素の不均一性、低バイオマスサンプル、宿主DNAのバックグラウンドによる交絡、ウイルス叢の分析のための標準的な計算ツールの欠如、および指数関数的に成長するウイルス叢データベースのために比較的数が限られています。しかし、限られた数のウイルス叢研究は、ウイルスが宿主、他のウイルス、および細菌との相互作用に応じて、人間の健康に有益または有害な影響を与える可能性があることを示唆しています。たとえば、クローン病におけるカウドウイルス(バクテリオファージ)の特定の拡大は、細菌の多様性の低下と関連していることが観察され、ウイルス叢が腸の炎症と細菌性腸内毒素症に寄与する可能性があるという考えを支持しています(Norman et al.,2015)。人間のウイルス叢の研究の多くは、ウイルス叢の変動と特定の病気との相関関係について報告していますが、メカニズムの理解は限られており、これらの変化したウイルス叢が実際に病気に寄与するのか、単にそれと相関するのかはわかりません。一方、一部のウイルスは防御的である可能性があります。たとえば、新たな証拠は、G型肝炎ウイルス(HGV)がヒト免疫不全ウイルス(HIV)関連疾患に対する保護効果をもたらす可能性があることを示しています(Lanteri et al.,2015)。健康と病気におけるヒトのウイルス叢はまだ揺籃期にあり、ウイルスが防御を提供できる正確なメカニズムについての知識は限られています。この現在のレビューの範囲を超えていますが、他の多くの最近のレビュー論文は、健康と病気におけるヒトウイルス叢の役割を調査しており、ヒトウイルス叢への関心の高まりとその幅広い影響を示しています(Zou et al.,2016; Neil and Cadwell,2018; Adiliaghdam and Jeffrey,2020)。 | |
| 人間のウイルス叢を片側に残し、より広い人間の胃腸マイクロバイオームに戻って これは、消化、ビタミン合成、免疫系の発達と維持など、人類生理学の複数の側面への貢献を考えると、人間の健康における重要な変数であることが繰り返し示されています(Olszak et al.,2012; Yatsunenko et al., 2012)。膣(Smith and Ravel,2017)、肺(Dickson and Huffnagle,2015)、皮膚(Byrd et al.,2018)など、他の解剖学的ニッチにおける微生物叢の人間の健康と病気への影響もかなりのものです。 | |
| 多種多様な要因が作用して個々のマイクロバイオームを形成し、潜在的に破壊します。これらは、遺伝学、加齢、生理学的パラメーター(pH、酸素レベル、運動性、ホルモン変動など)などの固有の要因(または自然)と食事、感染症、投薬などの外的要因(または育成/曝露)に大きく分けることができます(Levy et al.,2017)。特定のマイクロバイオームの状態が感染や病気に対する感受性を高める可能性があることを考えると(Mcburney et al.,2019)、プロバイオティクス(「適切な量で投与されると、宿主に健康上の利益をもたらす生きた微生物」)の有用性への関心が高まっています(Hill et al.,2014)、これはマイクロバイオームを有益に変化させて抗ウイルス免疫を強化する可能性を提供します。 | |
| このレビューは3つのセクションに分かれています。 1)マイクロバイオーム、宿主免疫系、ウイルス性病原体間の相互作用、 2)さまざまな曝露やライフイベント(食事、投薬、老化など)がマイクロバイオームを破壊し、ウイルス免疫にどのように影響するか、 3)プロバイオティクスによるマイクロバイオームの操作が、さまざまなウイルス感染をどのように予防または改善するか。 | |
マイクロバイオームとプロバイオティクスの抗ウイルスメカニズム |
|
細菌、ウイルス、宿主の生理機能の相互作用は複雑であり、まだ学ぶことがたくさんあります。それにもかかわらず、ウイルス性病原体に対する宿主の防御に対する共生生物とプロバイオティクス生物の両方の魅力的な貢献を明らかにする証拠が増え始めています(Li N et al.,2019)。ウイルスが粘膜表面(膣、呼吸器、消化管など)にさらされると、粘液層、自然免疫防御、獲得免疫防御という3つの幅広い防御線が克服されます(Kumamoto and Iwasaki, 2012)。証拠は、さまざまな共生細菌およびプロバイオティクス細菌がこれらの防御線のそれぞれに影響を及ぼし、さまざまなウイルス感染に重要な関連があることを示唆しています。責任のある抗ウイルスメカニズム(図1に要約)は、直接的および間接的の両方であり、次のものが含まれます。 1)強化された粘膜バリア機能(Lieleg et al.,2012; Schroeder,2019)、 2)抗ウイルス抗菌ペプチド(AMP)の分泌(Torres et al.,2013; Quintana et al.,、2014);バクテリオシン、 3)宿主細胞へのウイルス付着の阻害(Botićetal.,2007; Su et al.,2013)、 4)抗ウイルス性の先天性および適応性の白血球機能の調節(De Vrese et al.,2005; Jounai et al.,2012)。 |
|
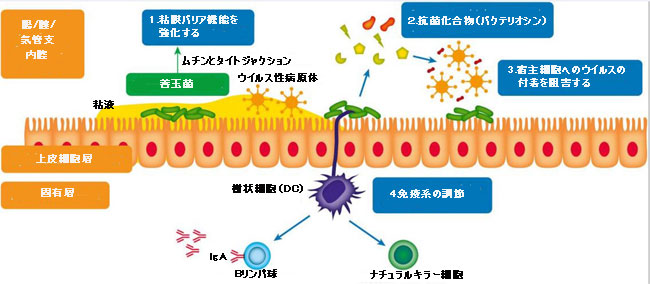 |
|
| 図1 抗ウイルスマイクロバイオームのメカニズム | |
| 1.強化された粘膜バリア機能:湿った上皮表面(消化管、膣、肺など)は、粘液と呼ばれる糖タンパク質を含む粘膜層で覆われています。これは、侵襲性の病原性微生物と宿主の上皮細胞との間に物理的なバリアを提供します。ムチン産生は胃腸マイクロバイオームの組成に影響され、抗ウイルス特性を持っているようです(Lieleg et al.,2012; Schroeder,2019)。多数の乳酸菌が密着結合を調節し、したがって正常な粘膜透過性を維持することが示されています。 | |
| 2.一部の細菌(例:バチルス属)によって生成される抗菌化合物(バクテリオシン)(例:サブチロシン)は殺ウイルス性です(Torres et al.,2013; Quintana et al.,2014)。 | |
| 3.さまざまな手段で宿主上皮細胞へのウイルスの付着を阻害します(Botićetal,2007; Su et al.,2013)。 | |
4.免疫系の調節:プロバイオティクスは形質細胞様樹状細胞を刺激してインターフェロンアルファ(IFN-a)を産生し、ウイルス感染に重要な役割を果たすナチュラルキラー(NK)細胞(自然免疫系)の細胞毒性活性を誘導します( Jounai et al.,2012)。 プロバイオティクス細菌株は、おそらく腸上皮細胞または免疫細胞の刺激による未知のメカニズムによって抗ウイルス免疫グロブリン(Bリンパ球によって産生される-適応免疫系)の産生を増強することも示されています(De Vrese et al.,2005)。 |
|
| 粘膜上皮は、病原性ウイルスや細菌に対する重要な障壁です。 この障壁の最初の構成要素は、人体のすべての湿った上皮表面(例えば、膣、消化管、および肺)を覆う多孔性生体高分子マトリックスである粘液です(Lieleg et al.,2012)。 粘液層はかつては受動的で厳密に宿主制御された構造であると考えられていましたが、最近の研究では、粘液層の形態と機能がマイクロバイオームの影響を受けることが示されています(Schroeder,2019)。 たとえば、消化管では、マイクロバイオームが胃腸粘液の糖タンパク質ネットワークの主成分であるムチン2をコードする遺伝子の発現を誘導します(Schroeder,2019)。 興味深いことに、ブタの胃ムチンは、ヒト乳頭腫ウイルス16型、メルケル細胞ポリオーマウイルス、A型インフルエンザウイルスなどのさまざまなウイルスによる上皮細胞の感染を防ぐことが示されています(Lieleg et al.,2012)。 さらに、膣マイクロバイオームの組成も頸膣粘液に影響を与えるようです。 Lactobacillus crispatusが優勢なマイクロバイオームは、Lactobacillus inersが優勢なマイクロバイオームで観察される急速な拡散とは対照的に、またはGardnerella vaginalisが大量に存在する場合に、HIV-1ビリオンの拡散を妨げることが示されています(Nunn et al.,2015 )。 | |
| 腸のマイクロバイオームが消化管バリア透過性に及ぼす影響、特に口語的に「リーキーガット」と呼ばれる透過性亢進との関連は、複数の疾患モデルで広く研究されています(Nagpal and Yadav,2017)。 上皮細胞間の傍細胞輸送を制御する多タンパク質複合体であるタイトジャンクションタンパク質の胃腸マイクロバイオームによる調節は、この場合、十分に立証されたメカニズムです(Al-Asmakh and Hedin,2015)。 さらに、Lacticaseibacillu scaseiやBifidobacterium adolescentisなどの一部の種は、感染時に密着結合の構造と機能を破壊するように作用するロタウイルス毒素NSP4の発現低下に関連する代謝物を生成することにより、バリア透過性を間接的に維持することがわかっています(Gonzalez-Ochoa et al., 2017)。 In vitroおよび動物研究の結果をヒト集団に変換することは困難ですが、さまざまな研究は、プロバイオティクスが粘膜バリア機能を改善できることを示唆しています(Bron et al.,2017)。 | |
| 抗菌ペプチドは、細菌からヒトに至るまで、事実上すべての生命体に共通する防御化合物です(Jenssen et al.,2006)。 バクテリオシンとしても知られる細菌の抗ウイルス抗菌ペプチド(AMP)は、すべての主要な細菌系統によって産生され、伝統的に重要なプロバイオティクス特性と見なされてきました(Dobson et al.,2012)。 バクテリオシンの細菌性病原体に対する抗菌活性は十分に立証されていますが(Dobson et al.,2012)、作用機序はそれほど注目されていませんが、抗ウイルス特性を持っているものもあります。 利用可能なデータから2つの行動様式が浮かび上がってきたようです。 一方では、バクテリオシンの中には、ウイルスがヒト細胞に侵入する前に抗ウイルス活性を示すものがあるようです。 この点で、ストレプトマイセスによって産生されるクラス1バクテリオシンであるデュラマイシンは、補助受容体TIM1をブロックすることにより、ジカウイルスの侵入を防ぐことがわかっています(Tabata et al.,2016)。 反対に、いくつかのバクテリオシンはウイルスの侵入を防ぎませんが、ウイルスサイクルの後期段階に干渉することにより、細胞変性効果とウイルス放出収量を減少させます。 たとえば、Lactobacillus delbrueckiiの5kDaバクテリオシンは、インフルエンザウイルス感染の初期段階には影響を与えませんが、感染細胞でのウイルスタンパク質の産生を減少させます(Serkedjieva et al.,2000)。 同様に、バチルス属の種によって産生されるバクテリオシンサブチロシンは、HSVタイプ1(Torres et al.,2013)とHSVタイプ2(Quintana et al.,2014)の両方の後期感染段階を破壊します。 特に、タンパク質合成段階までのウイルス複製は影響を受けませんが、ウイルス糖タンパク質gDの細胞内局在は変化します。 逆に、Enterococcus faecium由来のエンテロシンCRL35は、糖タンパク質gD合成を低下させることがわかったため(Wachsman et al.,2003)、サブチロシンと比較して、HSVの感染サイクルのわずかに早い段階を混乱させるようです。 | |
| ウイルス感染の最初の段階では、ビリオンが宿主細胞に付着します(Doms,2016)。 Lactobacillus (sensu lato) とBifidobacteriumの菌株は、細胞培養モデルにおいて、おそらく立体障害によって、水疱性口内炎ウイルスの細胞への付着と侵入を妨げることが示されています(Botićetal.,2007)。 さらに、細胞壁にCD4受容体を発現するLactobacillus spp(sensulato)は、HIV-1偽ウイルスに結合して捕捉できるため、CD4 +細胞の付着を防ぎ、ウイルス感染を減らすことができます(Su et al.,2013)。 | |
| 自然免疫系とは、ウイルス抗原への曝露に反応する非特異的で即時の防御機構を指します。 このシステムは、物理的障壁(例、上皮細胞表面)、細胞受容体(例、トール様受容体(TLR))、抗菌ペプチド(例、ディフェンシン)、および生来の白血球(例、ナチュラルキラー細胞(NK)および食細胞)で構成されます。 上皮細胞、マクロファージ、樹状細胞(DC)は、粘膜環境を継続的にサンプリングし、パターン認識受容体(PRR)を介して侵入ウイルスの存在を検出します(Holt and Strickland,)。 形質細胞様樹状細胞(pDC)は、ウイルスと細菌の両方を認識する特殊な免疫細胞であり、インターフェロンアルファ(IFN-a)の産生を介してナチュラルキラー細胞の細胞毒性活性を誘導する上で重要な役割を果たします。 インターフェロンは、ウイルスの核酸の転写を妨害することによってウイルスの複製を阻害するサイトカインです。 前臨床試験では、pDCによるLactococcus lactis JCM 5805株の食作用が、TLR9 / MyD88シグナル伝達を介してインターフェロン産生を有意に刺激しました(Jounai et al.,2012)。 ヒトでは、ナチュラルキラー細胞がウイルス感染に重要な役割を果たすことが認識されています。 さまざまな菌株と投与量を用いた6つのランダム化プラセボ対照臨床試験(RCT)の最近のレビューでは、プロバイオティクスが健康な高齢者のナチュラルキラー細胞活性を有意に増加させると結論付けました(Gui et al.,2020)。 しかしながら、著者らは、研究間の異質性と関与する参加者の数が少ないことを考えると、この結論は決定的なものではないことに注意した。 | |
| 適応免疫とは、抗体産生B細胞やCD4(ヘルパー)型およびCD8(細胞傷害性)型のT細胞など、病原体特異的記憶を産生する能力を持つ、遅延しているが病原体特異性の高い防御メカニズムを指します。 たとえば、プロバイオティクス株 L. rhamnosus GG(ATCC53103)は、腸内の上皮細胞にシグナルを送り、B細胞を刺激してIgA抗体を産生させる可溶性因子Msp2(またはp40)タンパク質を産生します(Wang Y. et al.,2017 )。 実際、L. rhamnosus GGを含むプロバイオティクス株は、動物モデルでIgAを増加させ(Wang Y. et al.,2017)、一部のウイルスワクチンの補助として投与した場合、ヒトでウイルス中和血清免疫グロブリンを増加させることが示されています(De Vrese et al., 2005) | |
マイクロバイオーム、腸内毒素症、およびウイルス感染症 |
|
腸内毒素症は、健康な個人に見られるものと比較して、居住者の共生コミュニティの構成の変化として広く定義されています(Petersen and Round,2014)。 この変化は、宿主と微生物の間の共生を混乱させ、潜在的に有害な結果をもたらすと考えられています。 健康なヒト腸内マイクロバイオームが構成するものは、簡単な答えのない質問であり、北米国際生命科学研究所のワーキンググループによって最近取り上げられたものです(Mcburney et al.,2019)。 いくつかの課題には、ヒトマイクロバイオームに見られる高度な個体内および個体間変動(たとえば、一卵性双生児は二卵性双生児よりもほとんど類似していません(Turnbaugh et al.,2010))、マイクロバイオームとホストの相互作用を定義および測定するための検証済みのバイオマーカーの欠如、そして、腸内毒素症が人類の生理学と病気の変化の原因または結果であるかどうかはまだ確立されていないという事実(Mcburney et al.,2019)が含まれます。 |
|
| 腸内毒素症は、潜在的な病原性種(ガンマプロテオバクテリアなど)の増加、有益であると考えられる生物( Bifidobacterium, Akkermansia あるいは Faecalibacterium 種)などの減少、アルファ多様性(特定の微生物叢における種の豊富さと均一性)の変化など、分類学的に頻繁に説明されます。 高いアルファ多様性は消化管の健康のマーカーですが、逆に膣のマイクロバイオームの腸内毒素症のマーカーです(Borgdorff et al.,2016)。 腸内毒素症は、有害な微生物由来の化合物(例えば、硫酸塩還元細菌によって生成される硫化水素)の生成などの機能用語で説明することもできます(Levy et al.,2017)。 実際、生態系の機能は、マイクロバイオームの特定の居住者の有無よりも重要である可能性があります(Mcburney et al.,2019)。 | |
| 多数の証拠が、マイクロバイオームの構造と機能の変化がウイルス免疫に影響を与えることを示しています。 このセクションでは、腸内毒素症とウイルス免疫の関係に関連するいくつかの重要なトピックについて説明します。これには、初期の生命因子、抗生物質への曝露、免疫老化と炎症、食事療法、炎症性疾患が含まれます。 | |
初期のマイクロバイオーム、免疫系、およびウイルス感染の間の相互作用 |
|
腸内毒素症が免疫系に与える影響は、おそらく生後1,000日間、すなわち受胎から子供の2歳の誕生日までの時間で最も重要です(Robertson et al.,2019)。 マイクロバイオームは、出産様式、食事、抗生物質への曝露など、宿主に関連するいくつかの環境要因の影響により、この期間中非常に動的です(Tamburini et al.,2016)。 幼少期には、一連の種を伴う生態学的な設置のプロセスがあり、最終的には比較的安定したコミュニティの確立につながります。 これらの初期の相互作用の変化は、粘膜および全身の免疫系の緊張を小児期、さらには成人期に設定するように思われます(Belkaid and Hand.2014)。 | |
| 新生児の最も重要な最初の微生物接種は、周産期に発生し、出産様式によって異なります。 帝王切開(CS)で出産した乳児は、一般的な皮膚( Staphylococcus, Streptococcus, または Propionibacteriaなど)(Tamburini et al.,2016)、病院環境に関連する日和見病原体(例:Enterococcus, Enterobacter, および Klebsiella 種)(Shao et al.,2019)が定着する傾向があります。経膣的に出産した乳児に見られるLactobacillus spp. (sensu lato)の初期の存在量とは対照的です。 これらの異なるコロニー形成パターンの程度は時間とともにあまり目立たなくなりますが、バクテロイデス門の多様性、および全体的な腸内細菌叢の多様性は、生後2年間の帝王切開出産児では低くなります(Jakobsson et al.,2014)。 この最初の違いは、細菌のコロニー形成が免疫系の発達と調節に必要であることを考えると、ウイルス免疫に重要な結果をもたらす可能性があります。 疫学研究では、帝王切開出産と、ウイルス感染のリスクの増加に関連する喘息などの慢性免疫障害(Sevelsted et al。、2015)との関連が示されています(Juhn,2014)。 したがって、出産様式、初期の細菌コロニー形成、および異常な免疫機能の間の関係は、因果関係がある場合、早期の予防的介入の可能性を示唆している。 プロバイオティクスによる初期の治療は、アトピー性皮膚炎などのいくつかのアトピー性疾患の発生率を減らすのに有益であることが示されていますが(Li L. et al.,2019)、これまでの喘息の結果はそれほど印象的ではありませんでした(Wei et al.,2020)。 | |
| 母乳で育てられた乳児は、人工乳で育てられた乳児と比較して、一貫してビフィズス菌のレベルが高く、全体的に多様性が低いため、早期の授乳は腸内細菌叢の組成に大きな影響を及ぼします(Milani et al.,2017)。 母乳は、ほとんどの新生児と乳児にとって最良の栄養源であると考えられています。 それは複雑な液体であり、微生物、炭水化物、および生物活性成分(IgAなど)の多様な配列を含んでいます。 Lactobacillus spp. (sensu lato) やBifidobacterium spp., など、人間の母乳に含まれる細菌種のいくつかは、最近発見された母子コミュニケーションの形態である腸-乳腺経路を介して母体の腸から移動する可能性があり、多くの呼吸器および消化管感染症から保護します(Le Doare et al.,2018)。 母乳には、構造的に異なる200を超えるヒト母乳オリゴ糖(HMO)も含まれています(Jost et al.,2015)。これは乳児が消化することはできませんが、ビフィズス菌やバクテロイデスなどの選択された属の成長を促進します(Marcobaletal.,2010)、消化管と免疫系の健康的な発達を促進すると考えられています。 ヒト母乳オリゴ糖は小腸で部分的に消化され、ほとんどが結腸に到達し、そこで発酵して短鎖脂肪酸(SCFA)と乳酸を生成します(Yu et al.,2013)。 これらの代謝物は、乳児の免疫系が成熟する間、いくつかの一般的な腸管ウイルスを不活化することが知られている酸性pHを生成します(Al Kassaa et al.,2014)。 このpH依存性の腸内細菌叢効果は、ウイルス感染性を抑制する腸内細菌叢の役割と、乳児の免疫系の「共同ヘルパー」としてのその役割に関する証拠を示しています(Li N. et al.,2019)。 | |
| 米国の6歳の子供たちの母乳育児と感染症の長期的な関連性に関する最近の前向き研究は、母乳育児が乳児期をはるかに超えて耳、喉、副鼻腔の感染症を予防する可能性があることを示唆しています(Li et al.,2014)。 証拠はまた、母乳育児が小児肥満の可能性を減らすことを示唆しており(Weng et al.,2012)、これは成人肥満のリスクを高めることが知られています。それ自体がいくつかのウイルス感染の危険因子です(Milner and Beck,2012)。 母乳育児、マイクロバイオーム、ウイルス免疫の間の相互関係の可能なメカニズムの説明は、最近、幼児アカゲザルで確認されました(Ardeshir et al.,2014)。 コミュニティの豊富さ、均一性、多様性は、人工乳で育てられた動物と比較して、母乳で育てられた動物のマイクロバイオームで大きく、TH1、細胞傷害性T、およびTH17細胞と正の相関がありました。 さらに、母乳で育てられた乳児は、Prevotella spp属菌とアラキドン酸が著しく豊富でした。 Prevotella sppは、リン脂質膜からアラキドン酸を放出することができるホスホリパーゼA2を発現します。大量のアラキドン酸を投与されたヒトの乳児は、ヒト免疫不全ウイルス+の母親からヒト免疫不全ウイルスに感染するリスクが低く、TH17集団の増加は、サル免疫不全ウイルス(Ardeshir)による感染に関連するウイルス負荷の減少と関連しているため、この発見はウイルス感染に特に関連しています(Ardeshir et al., 2014)。 乳児期における腸内細菌叢のこの重要な役割のもう1つの非常に最近の例は、腸内細菌叢組成とロタウイルスワクチンRotarixへの反応の関連性の調査です(Harris et al.,2017)。 この研究では、78人のガーナ人の乳児におけるRotarix®および糞便細菌叢に対する血清IgA抗体反応を分析し、結果として得られた39人の反応者と非反応者のガーナのペアをオランダの乳児と比較しました。 全体的なマイクロバイオームの組成は、ワクチン接種に対する反応者と非反応者の間で有意に異なり、ガーナの反応者は非反応者よりも健康なオランダの乳児に類似していた。 Rotarix®への反応は、 Streptococcus bovisの存在量の増加と相関し、反応の欠如は、Bacteroidetes門の豊富さ、特にBacteroidesおよび Prevotella属の種に関連するいくつかの細菌と相関しています。 この研究は、ワクチン接種後の乳児における腸内細菌叢の組成とセロコンバージョン率(訳者注:ワクチンの有効性を判断するために使用される方法の1つ)との相関関係を浮き彫りにしました。 最後に、腸内細菌叢は、腸内細菌叢が宿主の免疫系の発達と特に生後数年の免疫において重要な役割を果たしていることを示唆する証拠が増えているために、感染と免疫の研究の最前線に来ました。 | |
抗生物質誘発性腸内毒素症とウイルス免疫 | |
多くの疫学研究は、初期の抗生物質への曝露が、喘息、湿疹、食物アレルギーなどの免疫関連の病気を発症するリスクを高める可能性があることを示唆しています(Tamburini et al.,2016)。 ヒトでの初期の抗生物質曝露も抗ウイルス免疫を損なうように見えますが、この関係は因果関係ではなく、抗ウイルス免疫に影響を与える遺伝的変異によって説明される可能性があります(Semic-Jusufagic et al.,2014)。 ノトバイオティック(無菌)の抗生物質処理マウスを使用した研究では、共生細菌の操作がリンパ組織の発達障害、免疫細胞の恒常性の調節不全、消化管の感染症や炎症性疾患に対する感受性の変化を引き起こすことが示されています(Abt et al., 2012)。 げっ歯類モデルでは、妊娠中および授乳中の母親への抗生物質投与により、母親とその子孫の腸内細菌叢に実質的な変化が生じました(Gonzalez-Perez et al.,2016)。 この研究の乳児マウスは、ウイルス感染後の死亡率の増加と加速を経験しました。 抗原特異的IFN-γ産生細胞傷害性CD8+T細胞は、致死量以下に感染した乳児マウスで減少することがわかりました。 さらに、感染していない乳児マウスのCD8 + T細胞は、in vitroでの活性化に続いてIFN-γ(抗ウイルス防御に重要なサイトカイン)産生を維持する能力の低下も示しました(Gonzalez-Perez et al.,2016)。 さらに、抗生物質で治療されたマウスは、先天性および適応性の抗ウイルス免疫応答の障害を示し、全身性リンパ球性脈絡髄膜炎ウイルスまたは粘膜インフルエンザウイルスへの曝露後にウイルスクリアランスを大幅に遅らせます(Abt et al.,2012)。 これは、共生細菌が、自然免疫系の活性化閾値と感度を較正する強壮信号を提供するように見えるためです。 Bradley et alは、腸内細菌叢が肺間質細胞に信号を送り、それらをIFNプライム状態に保ち、ウイルスのライフサイクルを早期に鈍らせることでインフルエンザウイルス感染から保護できることを実証しました(Bradley et al.,2019)。 この研究での経口抗生物質治療は、げっ歯類におけるこの抗ウイルスメカニズムを損ない、したがってウイルス感染に対する感受性を高めることが示されました。興味深いことに、この抗生物質によって誘発された障害は、対照マウスからの糞便移植で逆転することが示されました(Bradley et al.,2019)。 さらなる研究では、経口抗生物質治療によって引き起こされた腸内毒素症は、膣粘膜の単純ヘルペスウイルス2型感染後の抗ウイルス免疫を直接損ないました(Oh et al.,2016)。 著者らは、共生細菌の枯渇が膣上皮からのIL-33の分泌に関連し、エフェクターT細胞の膣組織への移動を阻止することによって局所的な抗ウイルス免疫を抑制し、それによってIFN-γの産生を阻害したことを報告しました。 |
|
加齢に伴う腸内毒素症とウイルス免疫 | |
| 私たちが年をとるにつれて、私たちの体の機能の多くは徐々に低下します。 これは、健康状態、ライフスタイル、食事の変化と密接に関連しています。 運動性と認知力のより明白な低下に加えて、腸内細菌叢の組成と活性および関連する免疫応答にも変化があります。 しばしば言及される糞便ビフィズス菌の減少に加えて、糞便細菌叢の組成における他の変化が観察されています。 クロストリジウムクラスターXIVaからの酪酸産生菌の減少、およびFaecalibacterium prausnitzii および Akkermansia muciniphilaなどの抗炎症菌の減少が報告されています(Mangiola et al.,2018)。 糞便中の微生物の多様性の維持が健康的な老化に有益であると考えられていることを考えると、これは重要です(Kim and Jazwinski,2018)。 老化、免疫老化に関連する免疫機能の低下は、ナチュラルキラー細胞と食作用活性の低下を伴う、自然免疫の障害につながります。 これにより、ウイルス感染やワクチン接種に対する反応が低下する可能性があります(Przemska-Kosicka et al.,2018)。 プロバイオティクスは、高齢者のナチュラルキラー細胞および食作用反応を改善することが報告されています(Miller et al.,2019)。 | |
| 免疫老化に特徴的な免疫系のいくつかの側面が減少しているにもかかわらず、高齢者ではしばしば「インフラメイジング」と呼ばれる炎症誘発性の状態も高くなっています(Kim and Jazwinski, 2018)。腸内細菌叢は短鎖脂肪酸(Bron et al.,2017)やトリプトファン代謝物(Rothhammer and Quintana,2019)の生成など、さまざまなメカニズムを通じて腸のバリアを適切に維持するために不可欠であるため、このプロセスの主要なプレーヤーであると考えられています。 腸のバリア機能の低下は、リポ多糖(LPS)などの炎症誘発性の細菌由来製品の体内への移行に関連しています(Mangiola et al.,2018)。 これは次に、インターロイキン(IL)-1、IL-1受容体拮抗薬タンパク質(IL-1RN)、IL-6、IL-8、IL-13、IL-18、C-反応性タンパク質(CRP)および腫瘍壊死因子(TNF)-α、および形質転換成長因子(TGF)-βなどの免疫調節サイトカインなどの循環炎症マーカーのレベルの上昇に関連しています(Ctoi et al.,2020)。 | |
| この慢性的な低レベルの炎症は、腸のバリアの透過性亢進を悪化または維持します。 これは、活性酸素種(ROS)の産生の増加と相まって、T細胞機能の低下に寄与し(Pinti et al.,2016)、したがって感染中のウイルスクリアランスの障害の可能性があります。 プロバイオティクスが腸のバリアをサポートできるという説得力のある証拠があり(Bron et al.,2017)、これは加齢に見られる炎症性緊張の増加を抑制し、ウイルス感染やワクチン接種に対する免疫応答を高める働きをする可能性があります。 しかし、そのような介入が臨床的に有効であることが明確に証明されていない(Aiello et al.,2019)。 | |
食事関連の腸内毒素症とウイルス免疫 | |
| 約600万年前のチンパンジーからの分岐は、腸内細菌の多様性の実質的な喪失を伴いました(Moeller,2017)。 植物ベースの食品から動物組織への初期の食事のシフトは、この細菌の衰弱を引き起こした可能性がありますが、これは過去約200年間(つまり、産業革命以降)に劇的に加速され、精製された小麦粉と砂糖が食事に広く導入されました(Adler et al.,2013)。 胃腸マイクロバイオームは哺乳類の宿主の免疫系と深く統合されているため、現代のライフスタイルの結果である多様性の急速かつ実質的な収縮は、感染症や免疫関連疾患のリスクを高める可能性があります(Moeller、2017)。 食事はマイクロバイオームプロファイルにかなりの短期的および長期的な影響を与えることが知られています(David et al.,2014)。 アフリカの田舎とヨーロッパの都市の子供たちの間でマイクロバイオームを比較する人間の研究は、前者が以下によって特徴付けられることを発見しました: I)より高い微生物の豊富さと生物多様性、 II)バクテロイデスの有意な濃縮とファーミキュテスの枯渇、 III)病原性の低いEnterobacteriaceae (Shigella and Escherichia属) およびIV)食物繊維消化のための遺伝子を含むPrevotella 属およびXylanibacter属からの細菌の特定の豊富さ。当然のことながら、かなり多くの短鎖脂肪酸と関連しています(De Filippo et al.,2010)。 加工食品や動物性脂肪が多く、植物由来の要素が少ない西洋型食生活は、マイクロバイオームを変化させ、炎症を促進し、健康に重要な影響を与える免疫に悪影響を与えることが示されています(Tilg and Moschen,2015)。 具体的には、食事に発酵性繊維が不足していると、腸内細菌叢による短鎖脂肪酸の生成が減少します。これは、適応免疫と自然免疫の両方で重要な役割を果たすことが知られています(Krautkramer et al.,2016)。 マウスでは、酢酸を産生するビフィズス菌の枯渇により、感染症への感受性が高まり、消化管の炎症が促進され(Tilg and Moschen,2015)、肥満とは無関係に高脂肪食が腸内マイクロバイオームの組成を変化させました(Hildebrandt,2009)。 さらに、げっ歯類モデルでは、経腸栄養(例、非経口栄養-静脈内栄養)を動物から奪うと、腸内細菌のコロニー形成が大幅に変化し(例、プロテオバクテリアのレベルが上昇し(Demehri et al.,2015))、特定の粘膜免疫に欠陥が生じます(例えば、分泌型IgAレベルの低下)。 これらの変化は、マウスの腸内細菌転座の増加と呼吸器系抗ウイルス防御の喪失に関連しています(Renegar et al。、2001)。 したがって、食物繊維の変化、および結果として生じる短鎖脂肪酸プロファイル、ならびにエネルギー供給の経路は、マイクロバイオームに影響を与え、ウイルス免疫に影響を与える可能性があります。 | |
腸内毒素症関連の疾患とウイルス免疫 | |
| ヒトマイクロバイオームの共生の崩壊は、炎症性腸疾患(IBD)、肥満、関節リウマチ(RA)、喘息などの多くの慢性疾患の罹患率の増加に関連しており、その一因となる可能性があります(Lloyd-Price et al.,2016)。 これらの疾患は、局所および全身性炎症の一貫した特徴を誘発する役割を果たすと考えられている重要なマイクロバイオームの変化を特徴としています(Hand et al.,2016)。 COVID-19のパンデミックと肥満によって最近強調されたように、おそらく当然のことながら、これらの慢性炎症性疾患ではウイルス感染のリスクが高くなる可能性があります(Finer et al.,2020)。 さらに、免疫機能障害や腸内毒素症を特徴とする関節リウマチなどの慢性疾患は、ウイルス感染の病態生理学への洞察を提供するため、臨床的および機械的に非常に興味深いものです(Schett et al.,2020)。 ウイルス免疫におけるマイクロバイオームの中心的な役割は、腸内毒素症の状況でのヒトワクチン接種研究で強調されています:抗生物質による腸内細菌叢の調節は、ロタウイルスワクチン接種に対する免疫応答を変更することが示されています(Harris et al.,2017)。 インフルエンザの場合、抗生物質によって誘発された腸内毒素症は、H1N1ワクチン接種に対する反応を損ない(Hagan et al.,2019)、炎症が増強されました。これは、TLR5を介した腸内細菌叢センシングに関連している可能性があります(Oh et al.,2014)。 興味深いことに、TLR5遺伝子の変異は、多くの炎症性疾患に関連しています(Leifer et al。、2014)。 | |
| ここでは、炎症、マイクロバイオーム、ウイルス感染の間の考えられる関連性をさらに説明するために、いくつかの慢性炎症性疾患について説明します。 | |
| 炎症性腸疾患(IBD) | |
| 潰瘍性大腸炎(UC)またはクローン病(CD)のいずれかの炎症性腸疾患(IBD)患者は、ウイルス感染のリスクが高いため、血清監視、ワクチン接種、抗感染療法などの特定の抗ウイルス治療が必要です。 このリスクの増加は、疾患の病態生理学と免疫調節薬の両方によって誘発される免疫学的障害によるものです(Marínet al.,2015; Magro et al.,2017)。 これは、炎症性腸疾患患者におけるB型肝炎(HBV)およびインフルエンザワクチンの免疫原性の低下に影響を与えると考えられており、その根底にある病態メカニズムはまだ解明されていません(Marínetal.,2015)。 調節不全の免疫腸内細菌叢のダイナミクスは、炎症性大腸炎の多因子病因において重要な役割を果たす可能性があります。 炎症性患者のマイクロバイオームは、健康な被験者と比較して、より大きな不安定性と減少した多様性を示す傾向があります(Halfvarson et al.,2017)。 いくつかの病原性微生物(例えば、侵襲性 Escherichia coli)の存在は、Tヘルパー1およびTヘルパー17細胞を活性化し、それによって粘膜損傷を誘発します(Sartor and Wu、2017)。 これにより、微生物由来の抗原の取り込みが増加し、生存可能な腸内微生物の転座が起こり、免疫応答の調節不全を促進し続けます。 炎症性大腸炎患者の胃腸メタゲノムも非炎症性大腸炎患者対照とは著しく異なり、酸化ストレスに対する耐性の増加と炎症マーカーとの相関を示しています(Franzosa et al.,2019)。 プロバイオティクスは、炎症性大腸炎患者における微生物叢関連の影響を軽減し、たとえば腸の腸バリアの完全性に作用することによって正常な免疫機能を回復するのに役立つオプションを提供する可能性があります(Sartor and Wu,2017)。 現在の臨床研究の結果は、嚢炎および軽度/中等度の潰瘍性大腸炎におけるプロバイオティクスの役割を示唆していますが、クローン病にはありません(Abraham and Quigley,2017)。 最近のメタアナリシスでは、潰瘍性大腸炎でのより確実な研究が求められています(Kaur et al.,2020)。 | |
| 肥満 | |
| 多くの研究が、肥満が2009年のインフルエンザA(H1N1)パンデミック株による感染からのより悪い結果の予測因子であると報告しています(Milner and Beck,2012)。 肥満はまた、より長いインフルエンザAウイルス排出(Maier et al.,2018)およびより重症のCOVID-19感染(Stefan et al.,2020)と関連しています。 さらに、肥満はワクチン接種に対する反応障害と関連していることを示す証拠があります(Milner and Beck,2012)。 横断研究では、A. muciniphila, F. prausnitzii, Methanobrevibacter smithii および Christensenellaceaeなどの特定の細菌集団は、太りすぎまたは代謝的に不健康な個体と比較して、痩せた個体でよりよく表されることが示されています(Stenman et al.,2016)。 豊富な研究により、肥満に見られる腸内毒素症は、ウイルス免疫の障害の原因となる可能性のある低悪性度の全身性炎症に関連していることが確認されています(Hartstra et al.,2015)。 興味深いことに、メタ分析では、特定の分類群(Bifidobacterium breve, B. longum Streptococcus thermophilus Lactobacillus acidophilus, L. delbrueckii, および Lacticaseibacillus casei)のプロバイオティクス菌株が、人体測定の結果や、代謝性疾患を伴う空腹時血糖値やインスリンなどの代謝リスク要因にプラスの影響を与える可能性があることが示されました(Koutnikova et al.,2019)。 糖の消化や吸収、短鎖脂肪酸の生成の増加など、摂取されたプロバイオティクスによって腸内で媒介される特定のメカニズムが関与している可能性があります(Panwar et al.,2014; Stenman et al.,2016)。 | |
| 喘息 | |
| 喘息は、幼児期に再発するウイルス感染によって部分的に開始されます。 喘息は抗ウイルス免疫が不十分であり、呼吸器ウイルスによって悪化が引き起こされます(Altman et al.,2020)。 その結果、喘息患者はSARS-CoV-2に感染しやすく、重度の感染症のリスクが高いと考えられています。 しかし、喘息が実際にCOVID-19合併症の危険因子であるかどうかは現在不明であり(Hosoki et al.,2020)、これまでの研究では予想される発生率の増加を決定的に観察していません。 喘息または併用療法のTh-2サイトカインプロファイルは、この明らかな矛盾を説明する可能性があります(Liu S et al.,2020)。 肥満の人に見られる関連性と一致して、 A. Muciniphila と F. Prausnitziiのレベルは、健康な対照と比較してアレルギー性喘息の子供の腸内細菌叢で減少しています(Demirci et al.,2019)。 人間の介入研究から得られる証拠は幼児期に限定されており、たとえば、シンバイオティクスはビフィズス菌の増加に関連する喘息にプラスの効果を示しています(Wopereis et al.,2014; Zhao et al.,2019)。 | |
| 関節リウマチ(RA) | |
| 関節リウマチ患者は、免疫系機能および免疫抑制薬の疾患関連障害に起因する一般集団と比較して、感染のリスクが高くなります(Listing et al.,2013; Favalli et al.,2020)。 特にウイルス感染に関しては、ウイルス性上気道病原体、単純ヘルペスウイルス、帯状疱疹ウイルスによる感染のリスクが高いことが研究で示されています(Widdifield et al.,2013; Arleevskaya et al.,2017)。 関節リウマチ患者の口腔および糞便マイクロバイオームのメタゲノムショットガンシーケンシングおよびメタゲノム全体の関連分析により、健康な対照と比較してグラム陽性菌の濃縮およびグラム陰性菌の枯渇が確認されましたが、全体的な豊かさと多様性はグループ間で同等でした(Zhang et al。、2015)。 疾患修飾性抗リウマチ薬(メトトレキサートなど)は、マイクロバイオームの特徴を健康な対照の特徴に部分的に回復させるようです(Zhang et al.,2015)。 関節リウマチ患者での4つのプロバイオティクス試験の最近のメタアナリシスでは、ある程度の改善が確認されましたが、データが不足しているため、さらに明確な研究が必要です(Aqaeinezhad Rudbane et al.,2018)。 | |
| 全体として、特定の慢性炎症性疾患ではウイルス感染のリスクが高くなりますが、この関連性は体系的ではなく、疾患やウイルス性病原体によって異なります。 腸内毒素症のパターンも炎症性疾患によって異なり、これらのマイクロバイオームの特徴をウイルス感染感受性に関連付けるメカニズムは現在不明です。 それにもかかわらず、炎症性の状況(例えば、プロバイオティクス、プレバイオティクス、シンバイオティクスなど)で腸と肺の両方のマイクロバイオームを維持または回復することを目的とした介入は合理的であり、急性および将来のウイルス感染に対する免疫応答を間接的に高めるのに役立つ可能性があります。 | |
プロバイオティクスとウイルス感染症 | |
| プロバイオティクスは、最も一般的にはLactobacillus (sensu lato) とBifidobacterium属の細菌種です。 このような有機体は、炎症性成分(感染性下痢、アトピー性皮膚炎、炎症性腸疾患、片頭痛など)を伴う状態の治療としてよく使用されますが、その真の機能はより微妙です。 炎症誘発性(Chiba et al.,2010)と抗炎症性(Plaza-diaz et al.,2013)の両方の反応を誘発することが示されているためです。 マイクロバイオームを介して効果を発揮する治療法(例、プロバイオティクス、プレバイオティクス、ポストバイオティクス、生存不能な微生物細胞、微生物画分、または細胞溶解物(Aguilar-Toaláetal.,2018))は広く利用可能であり、多くの異なる診断のために長い間使用されてきました。 | |
| この最後のセクションでは、以下を含むいくつかの重要なウイルス感染症、すなわちウイルス性胃腸炎、ウイルス性肝炎、ヒト免疫不全ウイルス(HIV)、ヒト乳頭腫ウイルス(HPV)、ウイルス性上気道感染症(URTI)、インフルエンザ感染症およびワクチン接種、SARS CoV-2感染症、および集中的な病院ケアに関連してプロバイオティクスの証拠を探ります。 | |
| 選択されたプロバイオティクス株がこれらの抗ウイルス活性を誘発するメカニズムは完全には理解されていません。ただし、これらのメカニズムの仮説は存在します。 内因性細菌叢と同様に、プロバイオティクスは粘膜バリア機能を改善し、ウイルス粒子がこの物理的バリアを通過してウイルス性疾患の際にこのバリアを維持する能力を制限する可能性があります(Bron et al.,2017)。 選択されたプロバイオティクス乳酸菌について直接的な殺ウイルス効果が観察されており、過酸化水素などの一次代謝産物や有機酸によるpHの低下がそのような効果をもたらします(Zabihollahi et al.,2012)。 また、いくつかのバクテリオシンは、それらのよく知られた抗菌活性に加えて、抗ウイルス活性を示すことが観察されています(Torres et al.,2013; Quintana et al.,2014)。 粘膜表面へのウイルスの付着は、感染の最初のステップです。 乳酸菌によって産生される細胞外多糖類は、例えば、アデノウイルスのこの段階を効果的に妨害することが示されている(Biliavska et al.,2019)。 また、他の代謝物や細菌細胞断片がウイルス結合を妨害する可能性があり(Martínet al.,2010)、Levilactobacillus brevis細胞壁断片が単純ヘルペス複製を阻害することが報告されています(Mastromarino et al.,2011)。 しかし、プロバイオティクスの最もよく文書化された抗ウイルス効果は、免疫系の調節です。 選択されたプロバイオティクスは、ナチュラルキラー細胞活性と細胞傷害性活性を増加させることが報告されています(Miller et al.,2019)。 また、IFN-γ、IL-2、IL-12、IL-18などの抗ウイルス性サイトカイン応答および改善された抗体応答が報告されています(Lehtoranta et al.,2020)。 これらおよび他のメカニズムの多くが並行しておよび/またはその後に機能することはありそうもないことではありません。 | |
ウイルス性胃腸炎とプロバイオティクス | |
| 20以上のウイルス、細菌、寄生虫が急性胃腸炎を引き起こすことが知られています。 世界中で、ロタウイルスは胃腸炎の最も一般的な原因であり、毎年45万人以上がロタウイルス感染に起因しています(Allen et al.,2010; Gonzalez-Ochoa et al.,2017)。 アストロウイルス、ノロウイルス、サポウイルス、およびアデノウイルスは、急性ウイルス性胃腸炎の他の重要な原因です。 | |
| ロタウイルスタンパク質NSP1およびNSP4は、ロタウイルス胃腸炎の病因に重要であることが示されています(Gonzalez-Ochoa et al.,2017)。 NSP1はINF産生を阻害し、炎症性サイトカインのダウンレギュレーションを引き起こすことが示されています(Gonzalez-Ochoa et al.,2017)。 NSP4タンパク質はウイルス毒素であり、腸粘膜の構造に影響を及ぼし(緊密な接合部複合体を標的とし)、細胞内および細胞外の相対的な電解質バランスに影響を及ぼし、最終的に分泌性下痢を引き起こします。 興味深いことに、プロバイオティクスは、腸細胞内でのロタウイルスの付着、浸透、複製を妨げることにより、ウイルス排出の減少につながる可能性があるという証拠があります(Rigo-Adrover et al.,2017)。 最近の研究では、Lacticaseibacillus caseiとBifidobacterium adolescentisの代謝物が、ウイルス毒素NSP4の発現低下と関連していることも示されています(Gonzalez-Ochoa et al.,2017)。 | |
| ロタウイルス胃腸炎の従来の管理は、嘔吐や下痢によって失われた液体や電解質を補充するための経口補水液(ORS)で構成されています。 亜鉛サプリメントは、経口補水を改善することが示されているため、世界保健機関によっても推奨されています。 1998年に最初のロタウイルスワクチンが認可されたにもかかわらず、世界的に広く普及するまでには至っていません(Offit,2018)。 ロタウイルスワクチンへの普遍的なアクセスは、このウイルスの世界的な重要性を減らすことを願っていますが、それまで、公衆衛生に対する効果的な治療戦略の重要性は残ります。 | |
| プロバイオティクスは下痢性疾患での使用の長い歴史があり、感染性下痢性疾患の管理におけるそれらの有効性を確立するために多くのうまく設計されたランダム化比較試験(RCT)があります。 2010年、Cochraneのグループは、合計8,014人の参加者を含む63件のランダム化比較試験の系統的レビューとメタアナリシスを発表しました(Allen et al.,2010)。 複数の異なる原因菌と複数の異なるタイプのプロバイオティクス治療を含むこの広範囲にわたる分析は、急性胃腸炎のプロバイオティクスの非常に有用な概要を提供します。 このメタアナリシスは、下痢症状の明確で統計的に有意な減少を示しました。プロバイオティクスは、プラセボと比較した場合、下痢症状の平均期間を24.76時間(95%CI 15.9 – 33.6時間)短縮しました。 さらに、2016年のレビューでは、選択されたプロバイオティクス菌株により、下痢と入院の両方の期間が約24時間短縮されたと結論付けられました(Vandenplas,2016年)。 これらのようなメタアナリシスには、著しく不均一な製剤に基づく総括的な結論の固有の弱点や、多くの場合未確認の、したがっておそらく変動する原因となる病原体など、いくつかの重要な制限があります。 しかし、これらの注意にもかかわらず、プロバイオティクスは治療ガイドラインの主流になり始めています。 欧州小児消化器・肝臓・栄養学会(ESPGHAN)は、2020年に、特定のプロバイオティクス菌株(Saccharomyces boulardiiやL. rhamnosus GGなど)が複数の臨床試験からの明確な証拠に基づいているというガイダンスを発表しました。 以前は健康だった乳児や小児の急性胃腸炎の補助療法として安全に使用できる可能性があります(Szajewska et al.,2020)。 これに加えて、カナダ小児科学会は、ウイルス性胃腸炎の場合に症状の持続期間を短縮するためにプロバイオティクスを使用することを推奨しています(Marchand et al.,2012)。 対照的に、米国消化器病学会は最近、乳児の急性胃腸炎に対するプロバイオティクスの効果を文書化した研究のほとんどが北米以外で行われたと主張して、そのような使用に反対するよう助言しました(Su G. et al.,2020)。 急性胃腸炎の管理におけるプロバイオティクス臨床研究は、他の適応症と比較して比較的進んでいますが、最も効果的な菌株、または菌株の組み合わせ、および投与量を理解するには、さらに明確な証拠が必要であることは明らかです。 | |
ウイルス性肝炎とプロバイオティクス | |
| B型肝炎(HepB)感染症は、特に発展途上国において、世界的な医療問題です。 ウイルスと宿主免疫系との間の病原性相互作用は、肝障害を引き起こし、潜在的に肝硬変および肝細胞癌を引き起こします(Wang J. et al.,2017)。 腸-肝臓軸を介した腸内細菌叢の腸内毒素症(Milosevic et al.,2019)は、慢性B型肝炎(CHB)に関連する炎症や病原性代謝物の蓄積を含む重度の肝不全への進行を促進することが示されています(Wang J.et al.,2017)。 B型肝炎感染に関連する腸内毒素症は、潜在的に病原性の細菌(例えば、一部の腸内細菌科)の増加と潜在的に有益な細菌すなわち、 Bifidobacterium種の減少を伴うようです。(Zeng et al.,2020)。 ビフィズス菌によって提供される保護メカニズムには、エンドトキシンレベル(胃腸および血漿)の低下、腸内細菌叢の組成調節、抗菌剤産生、胃腸バリア機能の強化、および局所および全身免疫の調節が含まれます(Zeng et al.,2020)。 この状況は、慢性B型肝炎患者の腸内毒素症を監視および変更するための可能な治療戦略を提供します。 In vitro細胞モデル(HepG2.2.15-統合されたB型肝炎 DNAを含み、HBsAgを分泌する)では、B.alescentis SPM0212の細胞抽出物はB型肝炎を阻害し、その抗ウイルスメカニズムはIFNを介した抗ウイルス応答の4つの主要なエフェクター経路の1つMxGTPase経路に関連しています(Lee D. K. et al.,2013)。 さらに、2010年に発表された研究では、B型肝炎ワクチン接種スケジュール(1価の2回投与+ 1回の投与の組み合わせ)に、B型肝炎プラセボに対してウイルス表面抗体(HBsAb)免疫グロブリンG産生の増強が証明されているプロバイオティクスが追加された(生後6か月間)(Soh et al.,2010)。 | |
| 慢性Bがた肝炎の場合と同様に、腸内細菌叢も慢性C型肝炎(CHC)の進行および関連する合併症に関連しているようです(Inoue et al.,2018)。 腸内細菌のアルファ多様性は、健康な個人と比較して慢性C型肝炎患者で有意に減少し、臨床病期の重症度と関連しています(Inoue et al.,2018)。 興味深いことに、特定の種であるStreptococcus salivariusは、慢性C型肝炎の進行に関連して劇的に増加することがわかりました(Inoue et al.,2018)。 | |
| 胃腸マイクロバイオームは、多くの医薬品の薬物動態および薬力学を代謝および調節することが知られています。 興味深いことに、実験的な非ヌクレオシドC型肝炎ウイルスNS5Bポリメラーゼ阻害剤(C型肝炎ウイルスの阻害剤)であるデレオブビルは、細菌の調節を示唆する糞便ホモジネートと比較して、ヒト肝ミクロソームとのインキュベーション後に異なる代謝プロファイルを示しました(McCabe et al.,2015)。 | |
| B型およびC型肝炎ウイルスによる持続感染は、進行性塊状線維症および肝硬変につながる肝障害を引き起こします。 肝硬変の進行は、腸透過性の増加、小腸細菌異常増殖、および細菌転座に関連しています。 この状況により、微生物由来の化合物(エタノール、アセトアルデヒド、リポ多糖など)が門脈循環を介して肝臓にアクセスし、肝臓の炎症、線維形成、そして最終的には肝硬変を促進する炎症性サイトカインの産生を刺激します(Wang et al.,2019)。 性格の変化、運動障害、意識レベルの低下を特徴とする肝性脳症(HE)は、肝硬変患者の30〜45%に発生する生命を脅かす合併症です。 肝硬変患者における肝性脳症の正確な根底にあるメカニズムは不明なままですが、高アンモニア血症と全身性炎症が重要な役割を果たしていると考えられています。 興味深いことに、研究により、肝硬変患者の胃腸マイクロバイオームにおいて、Alcaligenaceae、Porphyromonadaceae、Veillonellaceae、Enterococcus、MegasphaeraおよびBurkholderiaが大幅に増加していることが示されています。 この分類学的変化は、高アンモニア血症と全身性炎症に関連しており、肝性脳症症状の悪化に寄与していました(Sung et al.,2019)。 非盲検では、ランダム化比較試験のプロバイオティクスが、肝硬変(ウイルス性肝炎に関連する36%)の患者の肝性脳症の二次予防に効果的であることがわかりました(Agrawal et al.,2012)。 さらに、前向き無作為化比較試験では、プロバイオティクスが肝硬変(ウイルス性肝炎関連の26.3%)患者の肝性脳症の一次予防にも有効であることがわかりました(Lunia et al.,2014)。 プロバイオティクスは、病原性の炎症誘発性のウレアーゼ産生生物を置き換えることによって作用し、肝性脳症の発症を防ぐ可能性があります。 | |
ヒト免疫不全ウイルス、膣マイクロバイオームおよびプロバイオティクス | |
| 膣の健康な微生物組成は、他の粘膜ニッチとは異なり、微生物の多様性が低いことを特徴とし、多くの場合、単一の Lactobacillus (sensu lato)種によって支配されています。 近年の分子技術の進歩により、生殖年齢の女性の膣マイクロバイオームは、5つの異なるコミュニティ状態タイプ(CSTs)に大まかに分類できることが明らかになりました(Smith and Ravel,2017)。 重要な共通点は、コミュニティ状態タイプが2つの明確なグループに分類されることです。 | |
| (i)Lactobacillus (sensu lato)優勢、Lactobacillus (sensu lato) の1つ以上の種が総コピー数またはシーケンシングリードの> 90%を占める場合(L. crispatus CST-I, L. gasseri CST-II, L. iners CST-III およびL. jensenii CST-V)、 | |
| (ii)非乳酸桿菌(non–Lactobacillus (sensu lato)) 優勢(CST-IV)、 Lactobacillus (sensu lato)は、厳格な通性嫌気性菌(Gardnerella属の種など)と30%未満の他の分類群のClostridialesの順序で多微生物混合物のを構成します(Smith and Ravel,2017)。 | |
| 膣内毒素症は、多様性の低い Lactobacillus (sensu lato) が優勢な微生物叢からの長期にわたる逸脱で構成されています。 無症候性の膣の炎症に関連する嫌気性多微生物性疾患である細菌性膣炎(BV)が最も一般的なタイプです(De Seta et al.,2019)。 細菌性膣炎は一般に抗生物質(メトロニダゾールまたはクリンダマイシン)で治療されますが、バイオフィルムに効果的に浸透できないため、有効性はしばしば最適ではなく、再発率は高くなります(Bradshaw and Brotman,2015)。 さらに、抗生物質の長期使用は、副作用や薬剤耐性のリスクを高めます。 最近の系統的レビューでは、34の臨床研究(van de Wijgert and Verwijs,2020)で、膣に適用されたLactobacillus(sensu lato)ベースのプロバイオティクスが膣のマイクロバイオームに与える影響を調べました。 著者らは、プロバイオティクスは細菌性膣炎の治療と予防に有望であると結論付けましたが、多くの研究の高い不均一性と次善の質が決定的な結論を妨げました。 プロバイオティクスが細菌性膣炎を改善するメカニズムには、病原体に対するコロニー形成抵抗性、炎症誘発性、バイオフィルム形成細菌(Gardnerella vaginalisなど)、健康的な低pH環境の維持(乳酸産生)、抗菌化合物の増加、頸膣粘膜免疫の調節などがあります( van de Wijgert and Verwijs、2020)。 膣のマイクロバイオームの状態は、ヒト免疫不全ウイルス(HIV)と重要な関連性があります。 細菌性膣炎は、女性のヒト免疫不全ウイルス感染リスクの増加(おそらく60%も)(Atashili et al.,2008)およびヒト免疫不全ウイルス排出の増加(Bolton et al。、2008)と関連しているため、細菌性膣炎治療はさまざまな手段によるヒト免疫不全ウイルス感染の減少に役立つ可能性があります。 さらに、CAPRISA(南アフリカのエイズ研究プログラムセンター)004試験では、抗レトロウイルス薬であるテノホビル(膣用ジェルとして使用)がヒト免疫不全ウイルス感染を全体で推定39%減少させることがわかりました(Karim,2010年)。 興味深いことに、研究者らは、テノホビルがLactobacillus (sensu lato) 優勢の女性でヒト免疫不全ウイルス発生率を61%(P = 0.013)減少させたが、非乳酸桿菌(non–Lactobacillus (sensu lato)) 優勢(CST-IV)の女性では18%(P = 0.644)しか減少させなかったことを確認しました(Klatt et al.,2017)。 G. vaginalisおよびその他の嫌気性細菌は、テノホビルを代謝することがわかったため、標的細胞がテノホビルを薬理学的に活性な状態に変換するよりも早く、テノホビルを枯渇させます(Klatt et al.,2017)。 まとめると、これらの調査結果は、ヒト免疫不全ウイルスの感染を減らすための新しいアプローチとして、膣内細菌叢を評価および調整することの潜在的な有用性を浮き彫りにしている。 | |
ヒトパピローマウイルス、膣マイクロバイオームおよびプロバイオティクス | |
| DNAウイルスの一種であるヒトパピローマウイルス(HPV)は、50歳までに性的に活発な女性の最大80%に感染すると推定されています(Moscicki,2005)。 ほとんどのヒトパピローマウイルス感染症は時間の経過とともに解消しますが、持続感染症は壊滅的な細胞周期の不安定性を引き起こし、最終的には浸潤癌、主に子宮頸部上皮内腫瘍および子宮頸部腺癌を引き起こす可能性があります。 それにもかかわらず、ヒトパピローマウイルスの存在だけでは癌の形成には不十分です。 上皮表面の完全性、粘膜分泌物、免疫調節などの粘膜表面に特有の個々の要因が、ヒトパピローマウイルスの持続性と癌への進行に関与している可能性があります(Pyeon et al.,2009; Schiffman et al.,2016)。 さらなる証拠は、頸膣細菌叢がウイルスの持続と退行に実質的な役割を果たし、したがってその後の疾患に重要な影響を与えることを示しています(Chang and Parsonnet,2010; Mitra et al.,2016)。 細菌性膣炎の特徴である慢性無症候性炎症と相まって、異生物性または多様性の高い膣内細菌叢は、ヒトパピローマウイルス感染の発生率、有病率、持続性の上昇と相関しています(Gillet et al.,2011; Guo et al.,2012)。 最近の観察横断研究は、特にL. iners CST-IIIおよび非乳酸桿菌(non–Lactobacillus (sensu lato)) 優勢(CST-IV)が、ヒトパピローマウイルス感染の存在および前浸潤性頸部疾患状態の発症と頻繁に関連しているという概念を支持しています(Lee JE et al.,2013; Brotman et al.,2014; Mitra et al.,2015)。 | |
| 特定の酵素および代謝産物の産生を通じて、細菌性膣炎関連微生物は、通常は基底ケラチノサイトへのヒトパピローマウイルスの侵入を阻害する頸部上皮バリア機能を損なうようです(Borgdorff et al.,2016)。 さらに、それらは持続的で生産的なウイルス感染を引き起こすいくつかの細胞経路に作用します(Hedges et al.,2006; Cheriyan et al.,2009)。 細菌性膣炎に関連するこれらの細菌性、粘膜性および免疫性の合併症はすべて、ヒトパピローマウイルス感染に対する感受性の増加および高悪性度の上皮内病変の発症をもたらす可能性があります。 膣のLactobacillus spp. (sensu lato)が細菌性膣炎関連微生物のコロニー形成を制限し、ひいては低pHの維持(Breshears et al.,2015)とバクテリオシンの生産(Stoyancheva et al.,2014)を通じて前述の有害なウイルス効果から保護するため、これは重要です。 膣内細菌叢の保護効果を詳しく調べると、Lactobacillus (sensu lato) プロファイルのタイプが極めて重要であることがわかります。 L. crispatusが優勢な膣内細菌叢は、高濃度のD-乳酸を生成します。これは、頸膣粘液の粘度を高め、ウイルス粒子の捕捉能力を高めることが最近示されています(Nunn et al.,2015)、 また、すべてのコミュニティ状態タイプ(CSTs)の中でウイルス感染の可能性が最も低い傾向があります(Borgdorff et al.,2014)。 | |
| プロバイオティクスは、ヒトパピローマウイルスとの関連で研究されており、生殖器のウイルスクリアランスと子宮頸部塗抹標本の質の向上を目的とした研究結果が得られています。 ヒトパピローマウイルスおよびPAP塗抹標本で低悪性度の扁平上皮内病変と診断された54人の女性を対象とした前向き対照研究を6か月間追跡した(Verhoevenet al。、2013)。 プロバイオティクス飲料を毎日摂取した後、介入群は対照群と比較して細胞学的異常の除去の可能性が2倍高かった(60対31%、P = 0.05)。 ヒトパピローマウイルスは、対照患者の19%に対して、プロバイオティクスユーザーの29%でクリアされました(P = 0.41)。 性器ハイリスクヒトパピローマウイルス(HR-HPV)感染症の女性121人を対象とした二重盲検(RCT)における、2つのプロバイオティクス株Lacticaseibacillus rhamnosusGR-1とLimosilactobacillus reuteriRC-14の3か月間の適用は、性器ハイリスクヒトパピローマウイルスクリアランスに影響を与えませんでした。 しかし、軽度の異常で不十分な子宮頸部スミアの発生率を低下させることができました(P = 0.017)(Ou et al.,2019)。 興味深いことに、膣プロバイオティクスの長期添加は、短期プロバイオティクス投与と比較して、細胞学的異常を軽減し(P = 0.041)、ヒトパピローマウイルスクリアランスを強化する(P = 0.044)優れた能力を持っていることが実証されています(Palma et al.,2018) 。 治療のアプローチや期間に関係なく、プロバイオティクスの機能的サポートと保護膣微生物叢の最終的な再確立は、ヒトパピローマウイルス感染とうまく戦うための重要な要素であるように思われます。 | |
ウイルス性上気道感染症およびプロバイオティクス | |
| 上気道は、鼻孔、鼻腔、口腔、扁桃腺、咽頭、喉頭で構成されています。 上気道感染症(URTI)は、これらの構造に影響を与える一連の病原性傷害を含む包括的な用語です。たとえば、風邪(主に鼻に影響を与える)、扁桃炎、咽頭炎、喉頭炎、急性中耳炎(中耳炎)、副鼻腔炎などです。 ウイルス上気道感染症は、ライノウイルス、呼吸器合胞体ウイルス、アデノウイルス、コロナウイルス、インフルエンザウイルス、パラインフルエンザウイルス、およびヒトメタニューモウイルスによって最も頻繁に引き起こされます(Bosch et al.,2013)。 急性上気道感染症は、人々が米国および世界で医療を求める最も一般的な理由です(Cherry et al.,2003)。 米国では毎年約10億の風邪が発生し、1人あたり年間平均2〜6回の風邪が発生します。 上気道感染症は一部の国で抗生物質使用全体の最大75%を占めており、このような感染症は主にウイルス性であるため、これらの貴重な医薬品の驚くべき誤用と、抗生物質耐性の危機を煽る1つの問題の規模を浮き彫りにしています(Fendrick et al.,2001; Hao et al.,2015)。 | |
| かなりの数の臨床試験が、上気道感染症を予防するためのプロバイオティクスの有効性を調べました。 大まかに言えば、プロバイオティクスは、局所および全身免疫の調節を通じて、胃腸感染の予防と同様の方法で気道感染を予防すると考えられています。 特定の効果には、末梢白血球の食細胞活性の増強、免疫グロブリン(IgA、IgG、およびIgM)の分泌の増加、およびサイトカイン(インターロイキン、TNF-α、インターフェロン-αなど)の産生の増加が含まれる場合があります(Hao et al.,2015)。 将来の研究は、臨床評価項目で見られる改善の原因となる、特定のプロバイオティクス菌株および菌株混合物の作用の根本的なメカニズムのより明確な理解を達成することを目的とすべきです(Ozen et al.,2015)。 | |
| 2015年のコクランレビューでは、合計3,720人の参加者が参加した上気道感染症予防のための12のプロバイオティクスのランダム化比較試験(RCT)が調査されました(Hao et al.,2015)。 試験には、ヨーロッパ、北アメリカ、南アメリカ、アジアのすべての年齢層(子供、大人、高齢者)のデータが含まれていました。 著者らは、急性上気道感染症のエピソードを経験している参加者の数を測定する場合、(少なくとも1つのエピソード:オッズ比(OR)0.53; 95%信頼区間(CI)0.37〜0.76、P値<0.001)、; 急性上気道感染症のエピソードの平均期間[平均差(MD)-1.89; 95%CI -2.03〜-1.75、P値<0.001];急性上気道感染症(OR 0.65; 95%CI 0.45から0.94)および風邪関連の学校欠席(OR 0.10; 95%CI 0.02から0.47)の抗生物質処方率の低下などのデータから、プロバイオティクスはプラセボよりも優れていると結論付けました。 | |
| すべてのメタアナリシスと同様に、プールされた有効性の結果の一般化には限界があります。 ただし、この分析に含まれる試験では、異種の介入(異なるプロバイオティクス株または株の組み合わせ)を利用していましたが、さまざまな投与量と期間で、菌株固有のメタアナリシスで確認されているように、少なくとも上気道感染症予防のためにプロバイオティクスを服用することの有望な治療効果があるようです(Poon et al。、2020)。 さらに、この同じコクランデータを利用して米国の人口に適用された経済モデリング研究は、プロバイオティクスの使用により、感染日数が5,400万日以上短縮され、抗生物質の投与が200万日以上回避され、400万日以上の休業日が回避されると結論付けました(Lenoir- Wijnkoop et al.,2019)。 コスト削減の観点から見たこのメリットは、米国の人口のみに換算すると、年間約14億米ドルに相当します。 どのプロバイオティクス株または株の組み合わせが免疫系の最も堅牢で信頼性の高い調節を達成し、最適な投与計画と介入のタイミングが何であるかを理解するための継続的な研究努力は、ウイルス上気道感染症のためのより効果的なプロバイオティクス療法の開発を可能にします。そして最終的にこれらの数字をさらに改善します。 | |
ヒトインフルエンザ感染症、ワクチン、およびプロバイオティクス | |
| ヒトインフルエンザウイルスA、B、およびCは、Orthomyxoviridae科のRNAウイルスです(Bouvier and Palese,2008)。 これらのウイルスは主に、上気道(鼻腔、口腔、咽頭、喉頭)から下気道(気管、気管支、細気管支、肺胞)まで走る呼吸器上皮細胞に付着して複製します(Kalil and Thoma,2019)。 インフルエンザは通常、軽度で合併症のない病気のみを引き起こし、ほとんどの人は医学的介入なしで回復します。 関連する疾患の重症度の主な決定要因は、ウイルスによって下気道が破られる程度であり、その危険因子には、極端な年齢(<5歳および> 65歳)、白人と併存する慢性心肺診断が含まれます(Kalil and Thomas,2019)。 この病気は世界中で発生し、北半球では、毎年のインフルエンザの流行(インフルエンザAおよびBウイルスによって引き起こされる)が秋と冬に発生し、人口の約5%〜15%に影響を及ぼします(Goeijenbier et al.,2017)。 インフルエンザの流行を引き起こすことが知られているのはインフルエンザAウイルスだけです。 最近の研究によると、インフルエンザに関連する季節性呼吸器死は、世界で毎年291,243~645,832人(10万人あたり4.0~8.8)であり、75歳以上の人々の中で最も高い死亡率(10万人あたり51.3~99.4人)を示しています。(Iuliano et al.,2018)。 | |
| 証拠は、消化管と呼吸器系が、それぞれのマイクロバイオームとともに、相互に通信し、影響を及ぼしていることを示唆しています。これは、腸-肺軸と広く呼ばれる関係です(図2を参照)(Dumas et al.,2018)。 | |
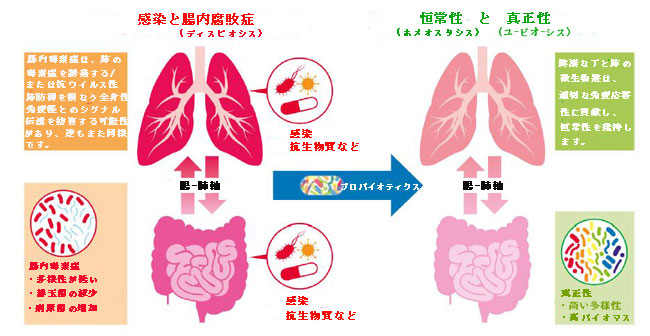 |
|
| 図2 ウイルス性呼吸器感染症の状況における腸-肺軸クロストーク | |
| 健康な腸内マイクロバイオームは、正常な免疫応答を維持することにより、呼吸器感染症に対する保護を提供します。 これは、多数の細菌由来の免疫調節化合物[例えば、リポ多糖(LPS)、ペプチドグリカン、および短鎖脂肪酸(SCFA)]による臓器間シグナル伝達によって達成される可能性があります。 たとえば、感染や抗生物質への曝露によって引き起こされる胃腸(GI)腸内毒素症は、これらのマイクロバイオームが生成する信号を(たとえば、血液を介して)変化させ、肺の抗ウイルス免疫応答を損なう可能性があります。 有益な細菌(すなわち、プロバイオティクス)の導入は、消化管および間接的に肺の効果的な免疫機能を強化または回復するのに役立つ可能性があります。 | |
| 特に興味深いのは、胃腸内マイクロバイオームの変化がウイルス感染に関連する肺免疫に影響を与えるように見えるという観察です(Dumas et al.,2018)。 インフルエンザ感染に関しては、この現象の正確な性質を解明する人間のデータはまだ生成されていません(Enaud et al.,2020)。 しかし、インフルエンザ感染げっ歯類モデルからの有望なデータは、プロバイオティクス株L. paracasei CNCM I-1518の摂取により、インフルエンザ感染に対する感受性が低下し、肺への炎症細胞浸潤が減少し、ウイルスクリアランスの速度が向上したことを明らかにしました(Belkacemetal.,2017)。 さらに、菌株から精製されたペプチドグリカン(主要な細胞壁成分)は、無傷の細胞の効果を部分的に再現することができました。 これらの化合物がウイルス感染を制御するメカニズムは、樹状細胞の肺への動員に関連していることが著者によって示唆されました。 追加のげっ歯類の研究では、肺組織のIFNシグネチャーを媒介するマイクロバイオームの役割が示されています。これは、初期のインフルエンザウイルス複製に抵抗性の環境を誘発し、初期のインフルエンザウイルス感染を鈍らせる可能性があります(Bradley et al.,2019)。 さらに、ネズミとヒトの両方の研究により、抗生物質が肺のIgA産生を減少させ、肺炎のリスクを高めるように作用することが明らかになりました(Libertucci and Young,2019)。 実際、動物実験では、胃腸内マイクロバイオームの抗生物質による根絶は、自然免疫応答と適応免疫応答の両方を損ない、CD4+およびCD8+ T細胞機能の障害を引き起こす可能性があることが示されています(Chen et al.,2017)。 さらに、げっ歯類における抗生物質誘発性腸内毒素症は、トール様受容体7(TLR7)およびNF-kB mRNAの発現を低下させ、抗ウイルス免疫の障害を引き起こしますが、これは、Bifidobacterium と Lactobacillus (sensu lato) のプロバイオティクス株の投与によって逆転しました(Wu et al.,2013)。 マウスモデルからヒト免疫系に直接外挿する前に注意する必要がありますが、この研究は、胃腸内マイクロバイオームと肺免疫系の間の興味深い相互作用を示しています。 | |
| インフルエンザの感染を減らすには、呼吸器症状のある人に社会的距離を置いて手指衛生を実践するように促すなどの公衆衛生対策が不可欠ですが、予防接種は依然として感染を防ぐための最も効果的な対策です。 ワクチン接種は、感染前に病原体を認識するように適応免疫系を刺激するように作用するため、その後の曝露が発生した場合に迅速な対応が可能になります。 ワクチンは一般的に2つの大きなカテゴリーすなわち弱毒生ワクチン(ロタウイルスワクチンなど)および不活化ワクチンに分けることができます。 これらにはいくつかのサブグループがあります。 不活化ワクチンはさらに次のように分類できます。 i)不活化ワクチン(例:ポリオ); ii)トキソイドワクチン、 ワクチンが病原体によって産生された毒素の不活化バージョン(例えば、ジフテリア)で作られている場合; iii)結合型ワクチン(例:髄膜炎c);およびiv)組換えワクチン(例えば、B型肝炎およびヒトパピローマウイルス)。 ワクチンは、伝染病の罹患率と死亡率の負担を大幅に軽減しました。 先進国では、天然痘を含むいくつかの感染症の完全な根絶はワクチン接種のおかげです。 さらに、ワクチン接種のおかげで、年間300万人の命が救われると推定されています(Toumi and Ricciardi,2015)。 栄養、性別、年齢、遺伝学、健康状態など、多くの要因がワクチンの有効性に影響を与えることを理解することが重要です(Vlasova et al.,2019)。 | |
| 残念ながら、特に高齢者にワクチン接種は感染からの適切な保護を提供できないことがよくあります。 ヨーロッパでは、リスクのある個人の44%だけが実際に毎年インフルエンザワクチン接種を受けています。 2009年にEU理事会勧告によって設定された75%の目標カバレッジ率(欧州連合理事会、2009年)からはほど遠い。 さらに、最近のレビューでは、65歳を超える成人のワクチンの有効性は37%であるのに対し、生産年齢の成人では51%であることが確認されています(Rondy et al.,2017)。 高齢者におけるこの効果の低下の理由は、しばしば免疫老化に起因します。 前に詳述したように、このプロセスには、いくつかの重要な免疫細胞タイプの減少が含まれます(たとえば、Tリンパ球、ナチュラルキラー細胞、樹状細胞数、およびB細胞の多様性の喪失)。 これにより、ワクチン接種および侵入する病原体に対する効果的な免疫応答を媒介する能力が低下し、両方とも感染症に対する感受性が高まります(Pera et al.,2015)。 | |
| インフルエンザワクチンやその他のワクチンを改善するための1つのアプローチは、免疫応答を高める物質アジュバントを含めることです(Tregoning et al.,2018)。 このような物質は通常、アルミニウム塩などのワクチンに添加されます。 これらの方針に沿って、免疫調節特性を特定する実質的な前臨床証拠に基づいて、ワクチン接種への反応を増強するためにプロバイオティクス(ヒト宿主に有益な効果を有する生きた微生物)を利用することに関心がありました(Rizzardini et al.,2012)。 これを支持して、ワクチン反応に対するプロバイオティクスの影響を調べた人間の臨床試験の最近の系統的レビューは、特定のプロバイオティクスがインフルエンザワクチン接種に対する免疫反応を増加させ、それがインフルエンザワクチン接種(血清変換)への反応が損なわれている高齢者の潜在的な利益を高めることを示唆する証拠を発見しました (Zimmermann and Curtis,2017)。 | |
| その後の系統的レビューとインフルエンザワクチン接種のアジュバントとしてプロバイオティクスを使用した臨床試験のメタアナリシスでは、赤血球凝集阻害(HI)抗体価が調べられました(Yeh et al.,2018)。 ヒト血清中のインフルエンザウイルス抗体の赤血球凝集阻害抗体力価試験は、ウイルス中和アッセイによって生成されたものと厳密に一致することが示されており、防御を予測します。 このメタアナリシスでは、著者らは、A / H1N1株、A / H3N2株、B株の両方のコントロールと比較して、プロバイオティクスグループで有意に高い赤血球凝集阻害力価を発見し、それぞれ20%、19.5%、13.6%増加しました。 このデータは、宿主の抗ウイルス免疫を増強するためのマイクロバイオーム操作の有望な役割を示唆しています。 | |
重症急性呼吸器症候群コロナウイルス2、マイクロバイオームおよびプロバイオティクス | |
| アンジオテンシン変換酵素2(ACE2)受容体は、重症急性呼吸器症候群コロナウイルス2(SARS-CoV-2)が結合してヒト細胞に侵入し、気道だけでなく胃腸上皮にも発現します。 初期の報告では、重症急性呼吸器症候群コロナウイルス2(SARS-CoV-2) RNAがかなりの割合の患者の便で検出されていたことが示されていました(Wu et al.,2020)。 生産的な感染は、in vitroで腸細胞オルガノイドで実証されています(Lamers et al.,2020)、 17件の研究の最近のメタアナリシスにより、重症急性呼吸器症候群コロナウイルス2 (SARS-CoV-2) mRNAが2019年のコロナウイルス病(COVID-19)患者のほぼ半数の便で検出されることが確認されました。 重症の場合や胃腸症状を示す場合の検出率は著しく高い(Gao et al.,2020)。 下痢は子供と大人で最も一般的な胃腸症状であり、平均期間は4.1±2.5日であるようですが、嘔吐は子供でより顕著であるようです(Tian et al.,2020)。 注目すべきことに、呼吸器サンプルが陰性になった後、一部の患者では便中のウイルスmRNA検出が維持されているようです(Wu et al.,2020)。 | |
| これらの観察は、病気の変数としての胃腸内マイクロバイオームの可能性を高めます。 実際、中国のコロナウイルス病(COVID-19)患者の初期の限定症例シリーズでは、Lactobacillus (sensu lato) とBifidobacteriumのメンバーが減少していることがわかりました(Xu et al.,2020)。 コロナウイルス病(COVID-19)患者を健康な対照と他の呼吸器感染症の入院患者の両方と比較した2つのその後の研究では、コロナウイルス病(COVID-19)患者の腸内細菌叢に重大な障害が見られました(Zuo et al.,2020; Gu et al.,2020)。 より具体的には、Actinomyces, Erysipelaclostridium, Streptococcus, Veillonella, Rothia, およびEnterobacterのような日和見病原体は、研究間のコロナウイルス病COVID-19)診断および/または重症度と関連していた。 逆に、Faecalibacterium と Anaerostipes, および Bifidobacterium のような有益な酪酸生成細菌は、両方の研究でコロナウイルス病(COVID-19)診断および/または重症度と逆相関していました。 さらに、マウス結腸でのアンジオテンシン変換酵素2(ACE2)発現をダウンレギュレートすることが知られている4つの特定の細菌(Bacteroides dorei、Bacteroides thetaiotaomicron、Bacteroides massiliensis、およびBacteroides ovatus)は、糞便中の重症急性呼吸器症候群コロナウイルス2 (SARS-CoV-2) mRNA負荷と逆相関しました(Zuo et al.,2020 )。 これらの研究の対照として他の状態で入院した患者の使用は、これらの発見に方法論的な強さを提供し、研究の1つは抗生物質の併用療法についても対照されました。 ただし、コロナウイルス病(COVID-19)の重症度における腸内細菌叢障害の原因となる役割はまだ確認されていません。 | |
| プロバイオティクスが提供する利点が何であれ、重症急性呼吸器症候群コロナウイルス2(SARS-CoV-2)感染に直接影響を与える可能性は低いです。 この点で、プロバイオティクスは、これらの特定の日和見細菌の増殖を阻害し、および/または有益な細菌の回復を促進することにより、一部のコロナウイルス病(COVID-19)症例で観察される腸内細菌叢障害の修正を促進する可能性があります。 さらに、プロバイオティクスは、免疫細胞との直接的なクロストークおよび/または腸透過性の低下によって免疫系の活動を高める可能性があります。これは、気道感染症に対するプロバイオティクスの有益な効果の背後にあるメカニズムの1つとして提案されています(Baud et al.,2020 )。 注目すべきことに、重症急性呼吸器症候群コロナウイルス(SARS-CoV)および重症急性呼吸器症候群コロナウイルス2 (SARS-CoV-2)細胞侵入はアンジオテンシン変換酵素2のダウンレギュレーションを引き起こし(Verdecchia et al.,2020)、アンジオテンシン変換酵素2欠損症はトリプトファン恒常性の障害をもたらすことが示されています(Hashimoto et al.,2012)。 腸内細菌叢の制御下にあるセロトニン、キヌレニン、およびインドール経路を横切るトリプトファン代謝(Agus et al。、2018)は、粘膜免疫および多数の全身効果に強い影響を及ぼします。 したがって、トリプトファン依存性経路は、コロナウイルス病(COVID-19)における特定の腸内細菌叢介入の標的を提供する可能性があります。 これに関して、健康な被験者へのプロバイオティクスL. plantarum DR7の投与は、プラセボと比較して、腸内細菌叢に特定の変化をもたらし、血清トリプトファン代謝経路を変化させることが最近示されました(Liu G et al.,2020)。 ただし、執筆時点では、コロナウイルス病(COVID-19)患者を対象としたランダム化プラセボ対照試験を完了したプロバイオティクス株はないため、それらの使用は実験的なままであることを強調する必要があります。 | |
クリティカルケアにおけるマイクロバイオーム、ウイルス感染症、およびプロバイオティクス | |
| 重症患者は集中治療室(ICU)で治療されます。集中治療室(CCU)または集中治療室(ITU)としても知られています。 このような患者は、集中治療室滞在中のある時点で少なくとも1つの臓器不全の有病率が高く(51%〜72%)、呼吸不全が最も蔓延しています(臓器不全の患者の87%を構成します)(Pedersenetal.,2017)。 臓器不全は、外部の臨床的介入なしに恒常性を維持できない程度の臓器機能障害として定義されます。 たとえば、呼吸不全の患者は、自分で呼吸することができないため、機械的人工呼吸が必要になる場合があります。 | |
| 急性呼吸窮迫症候群(ARDS)は、肺が重度の広範囲にわたる損傷に苦しんでいる状態であり、最も一般的には敗血症が原因で、進行性の呼吸不全を引き起こします。 肺炎は敗血症の最も一般的な原因です(Dickson,2017年)。 急性呼吸窮迫症候群(ARDS)は、重症急性呼吸器症候群(SARS)とH1N1インフルエンザウイルスの流行によって引き起こされた肺炎の最も致命的な合併症でした(Hendrickson and Matthay,2013)。 インフルエンザ感染の最も恐れられている結果の1つは、二次細菌感染です(これは、別の病原体、通常はウイルスによる感染中または感染後に発生する細菌感染です)(Morris et al.,2017) 罹患率と死亡率が大幅に増加したため、結果として急性呼吸窮迫症候群(ARDS)が発生しました。 呼吸不全は、慢性気道疾患(喘息、慢性閉塞性肺疾患、嚢胞性線維症など)の悪化によっても引き起こされる可能性があり、細菌はその病因において物議を醸す役割を果たしますが、ウイルスは明白な沈殿物です(Dickson,2017年)。 興味深いことに、インフルエンザAウイルスH3N2に感染したマウスによるプロバイオティクス株(Lactocaseibacillus paracasei CNCM I-1518)の経口摂取は、髄膜炎菌による二次細菌感染の負担を軽減することが示されました(Belkacem et al.,2018)。 | |
| 最近の研究では、集中治療室で治療された患者の糞便マイクロバイオームの実質的な不均衡(すなわち、腸内毒素症-特定の生態学的ニッチにおけるそれらの種の機能的能力に存在するいずれかの種の不均衡)が示されています(Wiersinga,2017)。 これは、重大な病気と集中治療の介入(抗生物質、プロトンポンプ阻害剤、食事の変更、概日リズムの乱れ、人工呼吸器のサポートなど)の両方の理解できる結果です。 集中治療室の介入は、身体に存在する微生物集団への影響に加えて、腸透過性(非ステロイド性抗炎症薬や非経口栄養など)と細菌の転座(腸粘膜から通常は無菌組織への細菌の移動)も増加させます(Dickson,2017)。この状況は、重篤な病気で発生する免疫調節不全を悪化させ、その結果、罹患率と死亡率の負担を増大させる可能性があります。しかしながら、重大な病気における腸バリア機能障害および腸内毒素症は、潜在的な治療標的を提供します。 多くの前臨床研究は、経口プロバイオティクスが腸のバリア機能と全身性免疫にプラスの影響を与えることができることを示しており、したがって、肺炎の場合、肺病原体の出現を抑制および除去する宿主の能力を高める可能性があります(Bron et al .,2017)(Forsythe,2014)。 確かに、重大な病気におけるマイクロバイオームの治療的増強のいくつかの有望な証拠が存在します(Schuijt et al.,2013)。 最近のレビューでは、人工呼吸器関連肺炎(VAP-機械的人工呼吸を48時間以上行った後に発生する肺の感染性炎症反応として定義)の発生率は、プロバイオティクスを服用している患者で安全に減少したと結論付けています(van Ruissen et al.,2019; Su M. et al.,2020)。 ただし、現在入手可能なデータの限界を考えると、さらに大規模な研究が必要です。 | |
| 急性呼吸窮迫症候群(ARDS)と敗血症の生存後でも、患者は集中治療室から退院してから数か月後に再入院するリスクが残念ながら高くなります(Dickson,2017)。 最近の出版物は、ほぼ11,000人の患者(アメリカ人、平均年齢77歳)の入院を調査し、いくつかの興味深い発見を報告しています。 1)重症敗血症の発生率は、入院後90日間(全原因入院の場合)で、この期間の前後のすべての期間よりも3.3倍高かった。 2)重症敗血症のための病院への再入院は、感染のために最初に入院した患者で有意に高かった(30%〜70%多い)(Prescott et al.,2015)。 感染症と抗生物質の両方の確立された結果である腸内毒素症は、この後者の関連を部分的に説明するかもしれません。 著者らは、入院中および入院後の腸内毒素症の期間は、免疫を調節するマイクロバイオームの通常の能力を損ない、病原性微生物の増加につながる可能性があり、それが圧倒的な宿主の炎症反応と相まって、重度の敗血症を引き起こす可能性があると仮定しています。 このいわゆる集中治療後症候群の根底にあるメカニズムはよくわかっていませんが、持続的に変化するマイクロバイオームの寄与を調査する必要があります(Dickson、2017)。 抗生物質の短期間のコースでさえ、マイクロバイオームの変化を数週間から数ヶ月持続させる可能性があることは十分に確立されており(Dethlefsen and Relman,2011)、プロバイオティクスは、抗菌薬による傷害後にマイクロバイオームを再確立するための有望な可能性を提供するようです(Mcfarland,2014 )。 マイクロバイオームが重大な病気の後にどれだけ迅速かつ完全に回復するか、そしてこのプロセスをプロバイオティクスで増強して臨床的に意味のある利益を実現できるかどうかを確立するには、さらなる研究が必要です。 | |
結論 | 21世紀における感染症の診断、予防、治療の進歩は、社会的、経済的、政治的に大きな利益をもたらしました。 しかし、気道感染症、ヒト免疫不全ウイルス、感染性胃腸炎だけでも、2030年には推定680万人の早期死亡(0〜69歳)が発生すると予測されています(Norheim et al.,2015)。 したがって、感染性伝染病は、警戒、監視、および新たな介入を必要とする人類に対する重大な脅威であり続けています。 細菌感染症を治療するための抗生物質と比較して、ウイルス感染症と戦うために開発された抗ウイルス薬は比較的少ない。 これは明らかに、満たされていない実質的な医療ニーズを表しており、単一のアプローチでは不十分です。 この例では、最近の研究は、さまざまなニッチにおけるヒトマイクロバイオームの状態がウイルスの病因に関連していることを示唆しています。 たとえば、腸内マイクロバイオームは、環境、遺伝、免疫のシグナルを同時に統合することで、宿主の代謝、免疫、感染への反応に影響を与えることができるシグナル伝達ハブとして機能しているようです(Thaiss et al.,2016)。 さらに、腸-肺および腸-肝臓の理解における最近の進歩(Milosevic et al.,2019)は、胃腸内マイクロバイオームの操作を通じて腸外臓器を標的とするウイルス性病原体に対する新しい防御手段を示唆しています(Dumas et al., 2018)。 膣のマイクロバイオームプロファイルは、赤血球凝集阻害ヒト免疫不全ウイルスやヒトパピローマウイルスなどのウイルス性病原体にも非常に関連しているようであり、したがって、これらの疾患の壊滅的な負担を軽減するための新しい標的を表しています。 |
| このレビューの最初の部分では、自然免疫系と適応免疫系の両方の要素を調節するなど、細菌株がウイルス感染から哺乳類の宿主を保護する可能性のあるいくつかの直接的および間接的なメカニズムの証拠を提示しました。 人間のこれらのメカニズムを明らかにし、利用することにおける継続的な進歩は、私たちが多くの悪名高いウイルス感染の伝染、罹患率および死亡率をさらに減らすことを可能にすることを願っています。 | |
| このレビューの第2部では、ウイルス感染に関連して研究を調査し、初期の生命因子(例、帝王切開および制限された授乳)(Tamburini et al.,2016)、抗生物質曝露(Dethlefsen and Relman,2011)、老化プロセス(例、免疫老化および炎症)(Mangiola et al.,2018)、不健康な慢性食事パターンおよびいくつかの炎症性疾患(例、炎症性腸疾患、肥満および関節リウマチ)を含む異種のストレッサーに対する哺乳類ホロビオントの微生物成分の感受性を実証しました。 これらの例でマイクロバイオームの操作を通じて抗ウイルス免疫を強化できるという見通しは魅力的ですが、因果関係は主に人間の被験者で証明されていないままです。 | |
| 3番目のセクションでは、胃腸管(ロタウイルス)、肝臓(BおよびC型肝炎)、頸膣上皮(ヒト免疫不全ウイルスおよびヒトパピロマウイルス)、呼吸器システム(ウイルス性上気道感染症病原体、特にインフルエンザに焦点を当てたもの、および重症急性呼吸器症候群コロナウイルス2(SARS-CoV-2)に関連するさまざまなウイルス性病原体の予防と管理におけるプロバイオティクスに関する現在のヒトの臨床的証拠をレビューしました。 これらの適応症の中で、プロバイオティクスはウイルス性下痢(Szajewska et al.,2020)および上気道感染症(Hao et al.,2015)の管理において最も広く研究されており、一般に、それらの有効性と安全性を裏付ける証拠は肯定的です。 ただし、このセクションで詳しく説明するウイルス感染の管理におけるプロバイオティクスの有効性の証拠には、さらに明確な研究が必要です。 | |
| 人間の生活は、季節性インフルエンザの流行や、コロナウイルス病(COVID-19)で現在生きているような散発的なパンデミックの脅威に常にさらされています。 社会的および衛生的対策を超えて、確実な保護を提供するために効果的なワクチンが必要であることは明らかですが、過去の経験は、そのような介入に対する望ましい免疫応答が期待外れである可能性があることを示しています(Rondy et al.,2017)。 したがって、プロバイオティクスが一部のワクチンに対する免疫応答を増強するためのアジュバントとして作用する可能性があるという証拠は有望であり、将来の大規模試験で調査する必要があります。 最後に、ウイルス性呼吸器病原体がそのような入院のかなりの数に直接的または間接的に関与していることを考慮して、病院の集中治療がマイクロバイオームに与える影響を調査しました。 この関係のダイナミクスを理解することで、これらの患者の罹患率と死亡率を低下させる可能性のある新しい治療手段が開かれます(van Ruissen et al.,2019; Su M. et al.,2020)。 プロバイオティクスの優れた安全性の記録と費用対効果の高い性質を考慮すると、最小限の臨床的に重要な利益を達成する見込みは、さらなる調査を価値のあるものにします。 | |
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
|
この文献は、Front Cell Infect Microbiol. 2020; 10: 596166.に掲載されたViral Infections, the Microbiome, and Probioticsを日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |